
Tłuszcz, mięśnie i dieta: dlaczego przy głodówkach tracimy nie tylko tłuszcz?
Poniżej tłumaczenie fragmentu publikacji pt. „A limit on the energy transfer rate from the human fat store in hypophagia”:
Ograniczenie szybkości transferu energii z ludzkiego zapasu tłuszczu w warunkach hipofagii
Na podstawie danych eksperymentalnych uzyskanych u osób niedożywionych, utrzymujących umiarkowaną aktywność, wyznaczono maksymalną szybkość transferu energii z zapasu tłuszczu człowieka w warunkach hipofagii [zmniejszone przyjmowanie pokarmu] i stwierdzono, że wynosi ona (290 ± 25) kJ/kg na dzień, a kaloriach (69 ± 5.9) kcal/kg na dzień. Restrykcja dietetyczna przekraczająca tę ograniczoną zdolność zmagazynowanego tłuszczu do kompensowania deficytu energetycznego prowadzi do natychmiastowego spadku beztłuszczowej masy ciała (ang. Fat Free Mass – FFM). W przypadku mniej nasilonego niedoboru energetycznego beztłuszczowa masa ciała [FFM] nie ulega redukcji. Rozwinięto koncepcję przejścia między tymi dwoma stanami dietetycznymi oraz zdefiniowano kryterium ich rozróżniania. Dla sytuacji, gdy zapas tłuszczu (ang. Fat Mass – FM) pracuje w trybie ograniczonego transferu energii, wyprowadzono ścisłe rozwiązanie matematyczne opisujące spadek beztłuszczowej masy ciała [FFM]. Rozwiązanie to ujawnia: składnik stanu ustalonego, zgodny z klasycznym rozumieniem, składnik wskazujący na powolny ubytek znacznej części beztłuszczowej masy ciała [FFM], łagodzony przez ograniczony transfer energii z zapasu tłuszczu [FM], oraz końcowy składnik pokazujący niechroniony, gwałtowny spadek pozostałej części beztłuszczowej masy ciała [FFM].
Wykazano, że średnia spoczynkowa przemiana materii u osób poddanych hipofagii spada liniowo w funkcji beztłuszczowej masy ciała, ze spadkiem równym (249 ± 25) kJ/kg na dzień. Wartość ta jest niezgodna z wynikami innych badaczy, którzy mierzyli tempo metabolizmu w różnych populacjach. Rozbieżność tłumaczy się indywidualnymi właściwościami metabolicznymi badanych osób, a nie charakterystykami większej populacji.
1. Wstęp
Popularne założenie w przypadku hipofagii głosi, że deficyty energetyczne są równoważone odpowiednim ubytkiem masy tłuszczowej (FM), co początkowo pozwala utrzymać stałość beztłuszczowej masy ciała (FFM). Zakłada się czasem, że stan ten będzie trwał aż do całkowitego wyczerpania zapasu tłuszczu (FM), po czym beztłuszczowa masa ciała [FFM] zacznie się zmniejszać. Choć paradygmat ten może wydawać się rozsądny, zostanie wykazane, że nie jest on prawdziwy w przypadku półgłodówki, gdzie beztłuszczowa masa ciała [FFM] spada od samego początku reżimu dietetycznego. Z danych eksperymentalnych wynika, że przy ciężkiej restrykcji dietetycznej zapas tłuszczu może dostarczyć tylko ograniczoną szybkość transferu energii do beztłuszczowej masy ciała, zmuszając do kompensowania deficytu kosztem ubytku beztłuszczowej masy ciała. Ograniczona zdolność masy tłuszczu do zaspokajania potrzeb energetycznych beztłuszczowej masy ciała może wynikać z limitowanych szybkościowo reakcji biochemicznych procesów transferu energii. Jeżeli jednak restrykcja dietetyczna nie jest głęboka, możliwe jest „oszczędzanie białka” przynajmniej do momentu, gdy masa tłuszczu spadnie do poziomu, przy którym jego ograniczona zdolność transferu energii zostaje zakwestionowana. W pracy omawia się oba przypadki, to jest „oszczędzania białka” i „braku oszczędzania białka” oraz rozważa przejście od pierwszego stanu do drugiego.
Aby wykazać natychmiastowy spadek beztłuszczowej masy ciała (FFM) podczas ciężkiej restrykcji dietetycznej, posłużono się danymi uzyskanymi w humanitarnym eksperymencie przeprowadzonym w czasie wojny na Uniwersytecie Minnesoty przez Keysa i in. (1950). Eksperyment ten będzie dalej określany jako eksperyment Minnesota. W ramach eksperymentu Minnesota 32 młodych mężczyzn – ochotników o statusie wojskowym – poddano półgłodówce w celu oceny optymalnych metod rehabilitacji do wykorzystania w leczeniu populacji niedożywionej w części ogarniętej wojną Europy. Dane z eksperymentu Minnesota są wykorzystane w niniejszej pracy z uwagi na długi okres kontrolowanej półgłodówki (24 tygodnie), wielokrotne pomiary masy tłuszczowej (FM) oraz wojskowo narzuconą i egzekwowaną dyscyplinę dietetyczną. Dla celów niniejszej analizy korzystne okazało się to, że średnia restrykcja energetyczna zastosowana w eksperymencie Minnesota – (6,56 ± 0,31) MJ/d – była niemal idealna do zilustrowania ograniczenia szybkości transferu energii z masy tłuszczowej.
W eksperymencie Minnesota masę tłuszczową mierzono trzykrotnie podczas 24-tygodniowego okresu półgłodówki za pomocą metod densytometrycznych, a wyniki skorygowano pod kątem nadmiaru płynów i minerałów. Skorygowane dane przedstawiono w formie tabelarycznej, bez oznaczenia niepewności eksperymentalnych. Autor niniejszej pracy posłużył się nieskorygowanymi danymi dotyczącymi masy tłuszczu i proporcjonalnie naniósł podane błędy bezpośrednio na zredukowane dane, które przedstawiono na ryc. 1. Słupki błędów na ryc. 1 odzwierciedlają błąd standardowy średniej i należy je uznać za minimalne, ponieważ nie obejmują nieznanych niepewności wprowadzonych przez proces korekcji. Dopasowanie metodą najmniejszych kwadratów do punktów eksperymentalnych dało wyrażenie:
f =9,51 exp [−(t/135)], (1)
gdzie f oznacza masę tłuszczu w kilogramach, a t czas w dniach. Równanie (1) przedstawiono na ryc. 1 ciągłą krzywą. Współczynnik korelacji dla tego dopasowania wyniósł 0,9991. Na ryc. 1 pokazano także popularną, niedynamiczną koncepcję (linia przerywana), zgodnie z którą stały deficyt energetyczny prowadzi do stałego tempa ubytku masy tłuszczowej. Krzywa kreskowo-kropkowana na ryc. 1 przedstawia dynamiczny spadek masy tłuszczowej przy założeniu braku ograniczeń w zdolności masy tłuszczowej do przekazywania dowolnej ilości energii wymaganej przez beztłuszczową masę ciała. Równanie opisujące tę krzywą zostanie wyprowadzone w dalszej części pracy i pokazuje, że masa tłuszczowa zostałaby wyczerpana po 98 dniach pół głodówki.
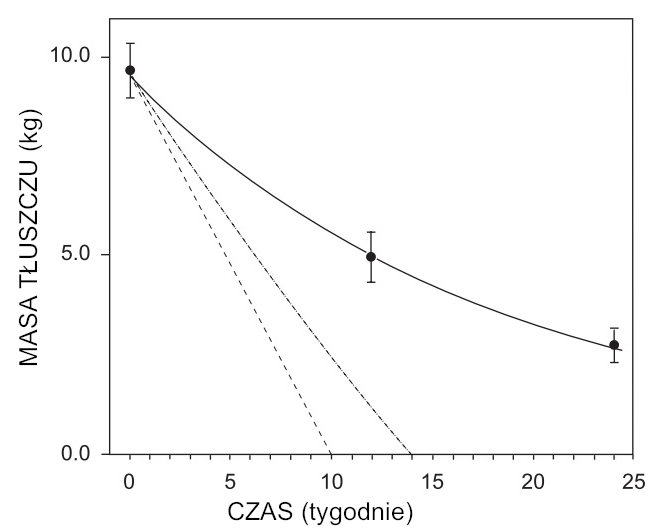
Rycina 1. Masa tkanki tłuszczowej (kg) w funkcji czasu (tygodnie). Ciągła krzywa to wykładnicze dopasowanie kwadratów trzech uśrednionych punktów danych eksperymentalnych. Linia przerywana przedstawia popularną koncepcję niedynamicznej utraty tkanki tłuszczowej, a krzywa przerywana przedstawia koncepcję dynamicznej nieograniczonej oszczędności beztłuszczowej masy ciała.
Punktowe odejmowanie wartości masy tłuszczowej uzyskanych z równania (1) od eksperymentalnych wartości całkowitej masy ciała dało wyniki przedstawione na ryc. 2 dla beztłuszczowej masy ciała (FFM) w trakcie pół głodówki oraz przez kilka tygodni przed rozpoczęciem restrykcji podaży energii z pożywienia. Z danych zaprezentowanych na ryc. 2 wynikają trzy obserwacje:
1. Następuje natychmiastowy spadek beztłuszczowej masy ciała.
2. Beztłuszczowa masa ciała osiąga wartość stałą w ostatnich 6 tygodniach półgłodówki. Ta stałość wynika z równowagi energetycznej pomiędzy nieznacznie zwiększoną podażą energii z pożywienia a obniżoną spoczynkową przemianą materii (ang. resting metabolic rate – RMR) oraz zmniejszonym wydatkiem energii na aktywność.
3. Rozrzut punktów eksperymentalnych względem hipotetycznej gładkiej krzywej mieści się w granicach błędu eksperymentalnego.
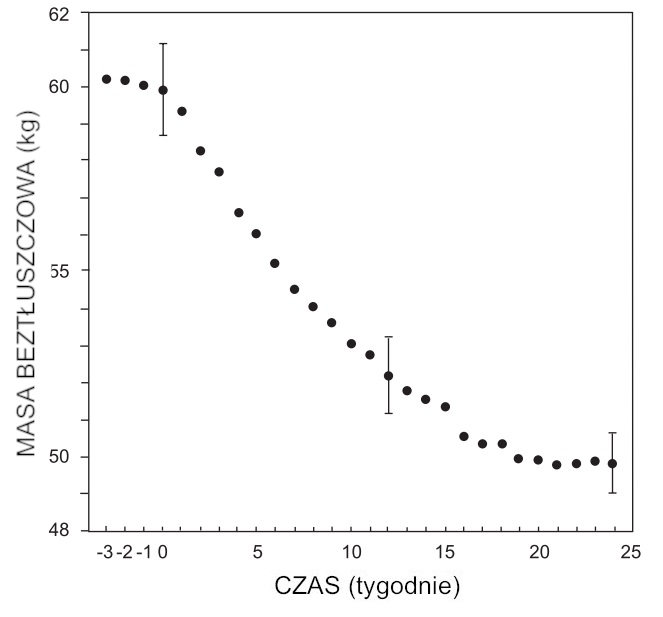
Rycina 2. Masa beztłuszczowa (kg) w funkcji czasu (tygodnie). Dane pochodzą z Keys i in. (1950).
Model energetyczny z dwoma rezerwuarami, zastosowany przez Alperta (1979), zostanie wykorzystany do opracowania teorii, która wyjaśnia główne cechy danych eksperymentalnych przedstawionych na ryc. 1 i 2. Model ten zakłada, że ciało składa się wyłącznie z masy tłuszczowej (FM) oraz beztłuszczowej masy ciała (FFM) i że energia jest magazynowana w obu tych rezerwuarach. W rzeczywistości ciało ma trzy oddzielne zasoby energii: cukier – glikogen, który znajduje się w mięśniach i wątrobie i związany jest z aktywnością mięśniową; białko, które występuje w wielu układach narządowych i postrzegane jest jako tkanka żywa; oraz masa tłuszczowa. W modelu włączamy glikogen i białko do beztłuszczowej masy ciała (FFM), wraz z wieloma innymi nieaktywnymi składnikami, takimi jak minerały kostne czy woda pozakomórkowa. Słowo nieaktywne w poprzednim zdaniu odnosi się do wszelkich składników ciała, które nie biorą bezpośredniego udziału w zużyciu tlenu. Fakt, że glikogen i białko [aminokwasy] zostały ujęte w ramach beztłuszczowej masy ciała, oznacza, że nie będziemy mogli oddzielić właściwości energetycznych tych dwóch magazynów energii. Główną przyczyną włączenia glikogenu i białka do jednej jednostki – beztłuszczowa masa ciała – jest to, że eksperymenty densytometryczne pozwalają rozróżniać i mierzyć jedynie masę tłuszczu oraz beztłuszczową masę ciała.
Masa tłuszczowa składa się z estrów glicerydowych i kwasów tłuszczowych i nie należy jej mylić z tkanką tłuszczową, której stanowi główny składnik. W sensie użytym w modelu masa tłuszczowa jest traktowana jako obciążenie zewnętrzne, fizjologicznie aktywne wyłącznie w procesie transferu energii do beztłuszczowej masy ciała. To właśnie beztłuszczowa masa ciała wchodzi w interakcję z tlenem.
Ponieważ beztłuszczowa masa ciała obejmuje białko, glikogen i wiele innych składników, rezygnujemy z używania określenia „oszczędzanie białka” (protein sparing) i zastępujemy je bardziej odpowiednim terminem: oszczędzanie beztłuszczowej masy ciała (FFM sparing).
(…)
5. Wnioski i spekulacje
Główna teza tej pracy brzmi: masa tłuszczowa (FM) jest w stanie przekazywać energię do beztłuszczowej masy ciała (FFM) z maksymalną szybkością (290 ± 25) kJ/kg na dzień. W realistycznych warunkach deficytu energetycznego rzeczywista szybkość transferu jest zmniejszona przez czynniki związane z aktywnością fizyczną. Wartość maksymalnej szybkości transferu została wyprowadzona na podstawie danych uzyskanych dla młodych, aktywnych mężczyzn badanych przez Keysa i in. (1950). Zastosowanie tych wyników nie zostało jednak bezpośrednio zweryfikowane w innych populacjach ani w odmiennych warunkach.
Badania eksperymentalne, których celem byłoby określenie wartości i rozkładu czynnika transferu energii, nie muszą być tak trudne ani czasochłonne jak oryginalna praca Keysa i in. (1950). Kolejne badania wymagałyby częstych pomiarów zarówno całkowitej masy ciała, jak i masy tłuszczowej w trakcie okresu dietetycznego, który zostałby zaprojektowany w taki sposób, aby początkowo oszczędzać beztłuszczową masę ciała. W momencie przejścia od oszczędzania beztłuszczowej masy ciała do braku oszczędzania beztłuszczowej masy ciała nie pojawią się oczywiste zmiany w masie tłuszczowej, natomiast powinny być zauważalne zmiany w beztłuszczowej masie ciała, które przejdą od wartości stałej do wartości malejących. Jeżeli takie zachowanie da się jednoznacznie zaobserwować, równanie σ fmin = | Δ| (18) może zostać użyte do obliczenia wartości realnego czynnika transferu energii.
W eksperymencie Minnesota zastosowano poważne ograniczenie dietetyczne, które natychmiast postawiło badanych w sytuacji braku oszczędzania beztłuszczowej masy ciała. W pewnym sensie było to korzystne, ponieważ pozwoliło na bezpośrednią interpretację zjawiska ograniczonego transferu energii. Gdyby wystąpiła sytuacja przejścia pomiędzy oszczędzaniem beztłuszczowej masy ciała, a brakiem oszczędzania beztłuszczowej masy ciała, jak pokazano na ryc. 5, interpretacja mogłaby nie być tak oczywista.
Jeśli koncepcja ograniczonej szybkości transferu energii z masy tłuszczowej jest prawdziwa, pojawia się ważne pytanie: czy taką szybkość można zwiększyć, aby umożliwić większą utratę masy tłuszczowej? Makroskopowa liniowa teoria energetyczna użyta w tej pracy daje niewielki wgląd w mechanizmy tego zjawiska. Bardzo prawdopodobne, że taka interwencja miałaby charakter farmakologiczny i wymagałaby jasnego zrozumienia odpowiednich mikroskopowych interakcji biochemicznych. Nasz model nie uwzględnia procesów nieliniowych, które mogłyby mieć znaczenie w nietypowych sytuacjach, ale które wciąż mogłyby być testowane za pomocą równania (18).
Niespodziewaną cechą maksymalnego czynnika transferu energii jest to, że jego wartość jest dość skromna. Alpert (1982) wykazał, że niska wartość σ’ lub σ jest bliska optymalnej, jeśli chodzi o wydłużenie okresu zmian w beztłuszczowej masie ciała. Taka optymalizacja pozwala na przedłużenie utrzymywania beztłuszczowej masy ciała nawet dwukrotnie w porównaniu do hipotetycznego przypadku nielimitowanego transferu energii.
Kolejna kwestia optymalizacyjna została przedstawiona w dokładnym rozwiązaniu dla beztłuszczowej masy ciała (FFM), pokazanym w równaniu (13). W tym równaniu ostatni składnik reprezentuje szybki spadek (28,6 dnia) niechronionej części beztłuszczowej masy ciała. W przypadku eksperymentu Minnesota początkowa wartość tego składnika wynosi 2,72 kg, czyli 22% całkowitej spodziewanej utraty beztłuszczowej masy ciała. W sytuacji optymalnej składnik ten powinien wynosić zero, ponieważ reprezentuje gwałtowną redukcję beztłuszczowej masy ciała.
Mimo że traktujemy beztłuszczową masę ciała jako pojedynczy byt energetyczny, w żaden sposób nie implikujemy, że beztłuszczowa masa ciała jest strukturą jednorodną. Możliwe, że chroniona część beztłuszczowej masy ciała dotyczy układów narządów życiowo istotnych, podczas gdy część niechroniona ma mniejsze znaczenie dla przeżycia. Jeśli matematycznie wymusimy, aby ostatni składnik równania (13) wynosił zero, a wszystkie inne zmienne pozostaną bez zmian, obliczamy, że maksymalna szybkość transferu energii z masy tłuszczowej wynosi 358 kJ/kg na dzień. Dla porównania, wartość zaobserwowana wynosi 290 kJ/kg na dzień.
Jak wskazano w równaniu (3), jedynym nieinwazyjnym sposobem, jaki ma jednostka do kontrolowania wielkości rezerw energetycznych, jest modyfikacja ilości spożywanej energii pokarmowej i/lub poziomu aktywności fizycznej. Dla osób żyjących w współczesnych społeczeństwach przemysłowych poziom aktywności jest głównie zdeterminowany przez zawód i styl życia i nie jest łatwo go zmienić. Jednak zawodowi sportowcy i oddani kulturyści byli w stanie kontrolować swoje rezerwy energetyczne poprzez ekstremalną aktywność.
Jednym z wyników omówionych w tej pracy jest to, że silnie ograniczony reżim dietetyczny ostatecznie prowadzi do znaczącej utraty nie tylko masy tłuszczowej, ale również beztłuszczowej masy ciała. Aby uniknąć utraty beztłuszczowej masy ciała, gdy masa tłuszczowa zostaje obciążona do granic swojej zdolności transferu energii, konieczne jest zakończenie lub złagodzenie drastycznego ograniczenia diety zanim dojdzie do takiej sytuacji. Poprzednia część pracy ilościowo sugeruje, jak można to zrobić. Popularne powiedzenie, że „jeśli trochę jest dobre, to więcej jest lepsze”, nie znajduje zastosowania w przypadku restrykcyjnych diet.
Choć głównym stanem dietetycznym omawianym w tej pracy jest niedojadanie (hipofagia), niektóre z tych samych rozważań dotyczą również przejadania się (Alpert, 1990). Równanie (3), które jest wyrazem zasady zachowania energii, powinno być prawdziwe w obu stanach dietetycznych. Podstawowa różnica polega na tym, że w przypadku przejadania się (hiperfagia) energia jest dodawana lub akumulowana zarówno w masy tłuszczowej, jak i w beztłuszczowej masie ciała. Głównym problemem modelu akrecji jest to, że nie jest jasne, w jaki sposób organizm decyduje o podziale nadmiarowej energii między te dwa magazyny.
Ciekawą cechę modelu akrecji można wyprowadzić z równania (3), zakładając, że albo masa tłuszczowa, albo beztłuszczowa masa ciała pozostaje stała, a wzrost drugiego izolowanego zasobu energetycznego jest analizowany. W ten sposób można pokazać, że przyrost masy tłuszczowej jest kilkukrotnie mniej kosztowny energetycznie niż taki sam przyrost beztłuszczowej masy ciała. Głównym powodem, dla którego wzrost beztłuszczowej masy ciała wymaga znacznie więcej energii, jest to, że spożywany pokarm musi dodatkowo dostarczać energii niezbędnej do zwiększonego podstawowego tempa metabolizmu (RMR). Znacząca różnica w kosztach energetycznych przy wzroście masy tłuszczowej i beztłuszczowej masy ciała empirycznie sugeruje, dlaczego niektórzy ludzie pozostają szczupli, a inni tyją podczas przejadania się. Sims i in. (1968) przeprowadzili długoterminowy eksperyment przejadania się na więźniach stanu Vermont i pokazali na wykresie słupkowym dla ośmiu badanych, że podział przyrostu masy między masą tłuszczową a beztłuszczową masą ciała był bardzo zróżnicowany.
Oczekuje się, że czas reakcji przejadania się w przypadku masy tłuszczowej będzie długi, a w przypadku beztłuszczowej masy ciała krótki, ponieważ ich izolowane wykładnicze równania wzrostu formalnie wykorzystują te same parametry, które zostały wprowadzone w sytuacji niedojadania. Ravussin i in. (1985) stwierdzili znaczny przyrost beztłuszczowej masy ciała już po 5 dniach przejadania się (Alpert, 1990).
Ostatecznie dochodzimy do wniosku, że w stanie półgłodówki beztłuszczowa masa ciała jest buforowana przed umiarkowanymi zmianami aktywności i jest w dużej mierze osłaniana przez energię transferowaną z masy tłuszczowej, która musi być odpowiednio duża, aby zapewnić pełną ochronę. W przypadku hiperfagii wnioskujemy, że masa tłuszczowa łatwo się powiększa. Wydaje się, że organizm ludzki jest dobrze przystosowany do środowiska ewolucyjnego typu „uczta-głód”. Problem, który obecnie rozwija się na całym świecie, polega na tym, że algorytm „uczta-głód” przestał działać. Najważniejsze pytanie brzmi: jak mamy radzić sobie z historycznie nową sytuacją permanentnej obfitości?
Źródło: J Theor Biol . 2005 Mar 7;233(1):1-13; A limit on the energy transfer rate from the human fat store in hypophagia
Zobacz na: Rzeźbienie Sylwetki – Drew Baye
Ten artykuł Cię wkurzy [insulina i glukagon]
Podstawy żywienia: Używanie widelca dla opornych
Tkanki tłuszczowej nie „spalamy”, tylko wydychamy
Większość utraty wagi [tłuszczu] następuje „poprzez oddychanie”
Przełom w dziedzinie tkanki tłuszczowej – dr Ellington Darden
Drew Baye: Na czym polega trening o wysokiej intensywności (HIT)?
Najnowsze komentarze