
Priorytet oksydacyjny, częstość posiłków i gospodarka energią uzyskiwaną z pożywienia i wydatkowaną na skutek wysiłku fizycznego: następstwa dla długości życia, otyłości i chorób kardiometabolicznych
Źródło: Metab Syndr Relat Disord. 2017 Feb 1;15(1):6–17; Oxidative Priority, Meal Frequency, and the Energy Economy of Food and Activity: Implications for Longevity, Obesity, and Cardiometabolic Disease
W większości współczesnych społeczeństw, stosunek wielu osób do żywności zmienił się fundamentalnie w porównaniu do poprzednich pokoleń. Ludzie odeszli od postrzegania żywności przede wszystkim jako pożywienia i obecnie poszukują produktów w oparciu o czyste walory smakowe lub konkretne potrzeby żywieniowe. Jednak kwestia tego jakie powinno być optymalne odżywianie ogółu populacji czy też konkretnych osób jest daleka od jasności. Wcześniej opisywaliśmy piramidę żywności jako sposób uporządkowania produktów opierający się na paradygmacie rosnącej gęstości energii, obecnie zaś rozszerzamy ten model, aby przewidzieć oddziaływanie priorytetu oksydacyjnego i zarówno gęstości składników odżywczych, jak i gęstości błonnika w odniesieniu do liczby spożytych kalorii. W połączeniu z częstością spożywania posiłków, zintegrowanym wydatkowaniem energii, priorytetem oksydacyjnym dotyczącym makroelementów oraz podziałem paliwa wyrażonym przez współczynnik oddechowy, nasz model oferuje także nowatorskie wyjaśnienie chronicznego przejadania się i przyczyny nadmiernego gromadzenia tkanki tłuszczowej. W niniejszym artykule analizujemy nie tylko metabolizm jako dynamiczny proces, podlegający wielu regulatorom, które wpływają na to, co się stanie z przyswojonymi kaloriami, ale omawiamy również kwestię w jaki sposób piramida żywności przewiduje priorytet oksydacyjny przyswojonego pożywienia oraz zapewnia konceptualny model zdrowego odżywiania wspierany badaniami dotyczącymi zdrowia i długości życia.

„Pomiędzy bodźcem a reakcją istnieje przestrzeń. W tej przestrzeni znajduje się nasza moc wyboru reakcji. W naszej reakcji leży nasz rozwój i nasza wolność” – Viktor E. Frankl
Wstęp
W ciągu zaledwie dwóch wieków przeszliśmy od odkrycia roli tlenu w procesie oddychania do badania złożonego, opartego na biologii molekularnej modelu oddychania komórkowego, regulacji hormonalnej i bioenergetyki w fizjologii i patofizjologii człowieka. Ponad stulecie temu żywność analizowano systematycznie pod kątem podstawowych składników energetycznych – białek, węglowodanów i tłuszczów – zaś biodostępność tej energii była określana ilościowo jako podstawa wymiany kalorii, której celem jest odżywianie. Wykorzystujące metodę kalorymetrii bezpośrednie i pośrednie doświadczenia z udziałem pojedynczych osób, w ramach zarówno długo-, jak i krótkookresowych badań w komorze metabolicznej, uwzględniających zarówno odżywianie, jak i wysiłek fizyczny, również dokładnie udokumentowały konserwację energii. Dlaczego więc popularne poradniki, artykuły naukowe i nagłówki w mediach wciąż kwestionują wiarygodność kalorii żywieniowej (z technicznego punktu widzenia kilokaloria [1000 kcal] lub ~4.2 kJ) w przewidywaniu deficytu energetycznego i nagromadzonego nadmiaru? I dlaczego, mimo że wiemy więcej o zarządzeniu energią niż kiedykolwiek wcześniej, wydaje się, że nasze społeczeństwo jest niezdolne do zarządzania skądinąd nieskomplikowaną równowagą energetyczną?
W krajach rozwiniętych stosunek wielu osób do żywności różni się fundamentalnie od podejścia poprzednich pokoleń. Nastąpiło przejście od postrzegania żywności jako pożywienia do zwracania uwagi na czyste walory smakowe lub konkretnych potrzeby żywieniowe (np. diety wysokobiałkowe, diety niskotłuszczowe, diety niskowęglowodanowe bądź ich połączenia). Jednakże daleko nam do wyjaśnienia jaki jest optymalny sposób odżywiania populacji ogólnej lub też konkretnych osób. I jak zrozumieć wszystkie zasady określonych modeli żywieniowych wraz z wszechobecnymi, sprzecznymi ze sobą przymiotnikami charakterystycznymi dla reklam, takimi jak świeży, organiczny, bezglutenowy, niskotłuszczowy, niskowęglowodanowy czy bez cukru?
Jak wspomnieliśmy wcześniej[1], piramida żywności przedstawia nową organizację żywności w oparciu o paradygmat rosnącej gęstości energii. Jednak piramidę żywności można dalej rozbudowywać, aby przewidzieć oddziaływanie priorytetu oksydacyjnego i zarówno gęstości składników odżywczych, jak i gęstości błonnika w odniesieniu do liczby spożytych kalorii. W połączeniu z częstością spożywania posiłków, zintegrowanym wydatkowaniem energii, priorytetem oksydacyjnym dotyczącym makroelementów oraz podziałem paliwa wyrażonym przez współczynnik oddechowy (RQ, ang. respiratory quotient), nasz model oferuje także nowatorskie wyjaśnienie chronicznego przejadania się i przyczyny nadmiernego gromadzenia tkanki tłuszczowej.
W niniejszej krótkiej analizie omówimy cztery główne punkty: (i) odżywianie nie jest sytuacją wyjątkową; (ii) piramida żywności przewiduje priorytet oksydacyjny; (iii) stan chronicznej sytości jest antytezą ograniczenia diety (DR, ang. dietary restriction) i opiera się korzystnemu oddziaływaniu DR na metabolizm, które wykazało badanie długości życia; i (iv) zmiany RQ indukowane ćwiczeniami zaburzają proces pozbywania się tkanki tłuszczowej i zapowiadają niezamierzoną kaskadę epizodów żywieniowych.
Przyswajanie kalorii, spalanie kalorii
Jedną z początkowych zasad metabolizmu człowieka jest to, że spożywaną żywność można określać ilościowo jako jednostki wymiany energii w oparciu o energetykę biologiczną i powiązane z tym ciepło odpadowe. Przez całą drugą połowę XIX wieku i na początku wieku XX badacze skrupulatnie pracowali nad zdefiniowaniem tej równowartości wymiany lub efektywności trawienia[2-6]. Spożyte pożywienie jest rozkładane na składniki, które są przechowywane, spalane lub usuwane w odchodach. Porównując dokładnie dane z kalorymetru bombowego dla różnych produktów żywnościowych z pomiarami składników takich jak ciepło i oddychanie w ciągu kilku godzin po posiłku, Carl von Voit, Max Rubner, Max Pettenkofer, Wilbur Atwater, Francis Benedict i inni obliczyli przeciętne konwersje energii dla białek, węglowodanów, tłuszczów i alkoholi.[5-10] Podczas gdy prawdą może być, że produkty izokaloryczne nie zawsze są izometaboliczne, spożycie kalorii z żywności minus rozproszone ciepło, oddychanie i te kalorie, które zostają usunięte jako odchody musi być równe zeru.
Chociaż może się wydawać, że niektóre kalorie dominują nad gromadzeniem się i utratą tkanki tłuszczowej, rozbieżność tę przypisuje się ogólnie upraszczaniu przeliczania energii. Z wyjątkiem rzadkich chorób genetycznych, otyłość w głównej mierze powoduje nadmierne spożycie żywności, a w mniejszym stopniu brak wysiłku fizycznego ze względu na tendencję do wzrostu zużycia kalorii w czasie ćwiczeń.[4,11] Jednakże zjawisko apetytu nabytego, które dominuje nad odruchowymi bodźcami żywieniowymi uznawane jest od czasów Hipokratesa12, zaś mechanizmy zaangażowane w doprowadzanie niektórych osób do przejadania się, a w przypadku innych do osiągnięcia naturalnej równowagi między głodem a wagą pozostają niewyjaśnione.[13,14] Otyłość nie jest powszechna w królestwie zwierząt z wyjątkiem zwierząt (np. domowych), które przekarmiamy lub wówczas, gdy dodatkowe kalorie są w sposób nienaturalny łatwo dostępne. Co więcej, w środowisku ubogim w kalorie, pragnienie przejadania się nie jest cechą doboru negatywnego i może faktycznie zapewniać konkretne korzyści. Zgodnie z poniższym opisem, tendencję do magazynowania części posiłku o największej gęstości energetycznej (tj. tłuszczów) można również wyjaśnić w kontekście przewagi adaptacyjnej.
Termodynamika przejadania się
Choć pod względem arytmetycznym wygodnie jest się skupić na dziennym nadmiarze lub niedoborze kalorii, dokładniejsze przedstawienie gromadzenia się kwasów tłuszczowych lub utleniania musi uwzględniać badanie na krótszej skali czasu konkretnego, wykorzystanego substratu lub składnika pokarmowego wraz ze wzrostem energii w stosunku do potrzeb podstawowych ze względu na dietę, aktywność oraz środowisko. Rycina 1 ilustruje dopływ i usuwanie kalorii w typowym dniu z wykorzystaniem godzinnych przerw, ze wskazanym czasem trwania, częstotliwością i częściowym pokrywaniem się głównych elementów energii. Dzienny całkowity wydatek energetyczny (ang. TEE, total energy expenditure) jest połączeniem podstawowej przemiany materii (ang. BMR, basal metabolic rate), aktywności, ćwiczeń, termogenezy indukowanej pożywieniem (ang. DIT, dietary-induced thermogenesis) i termogenezy indukowanej środowiskiem (ang. EIT, environmentally-induced thermogenesis). Na drodze eksperymentu BMR mierzona jest na czczo po obudzeniu się w ciemnym pokoju. Jednak podstawowa przemiana materii [BMR] jest często stosowana jako synonim przemiany materii w stanie spoczynku (ang. RMR, resting metabolic rate), którą zazwyczaj mierzy się mniej rygorystycznie i która wymaga jedynie stanu na czczo oraz braku aktywności fizycznej zwykle przez 4 godziny przed badaniem.[15] Badania doświadczalne zarówno dla podstawowej przemiany materii [BRM], jak i przemiany materii w stanie spoczynku [RMR] trwają zwykle 15-20 minut, zaś wyniki ekstrapolowały w ciągu 24 godzin.[15]
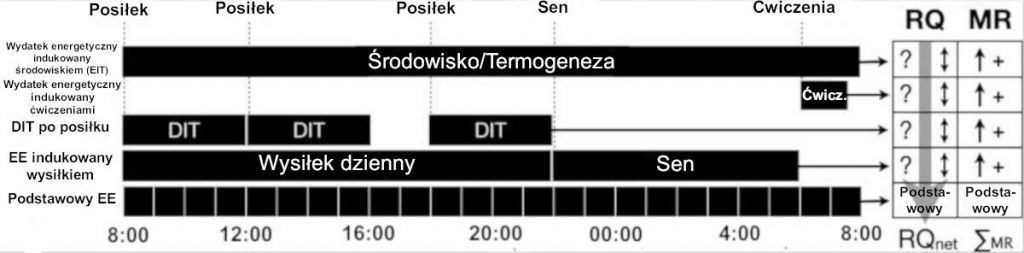
RYC. 1: Zmiana BMR i związany z nią wpływ na utlenianie tłuszczów. Dla każdej godzinnej przerwy w ciągu 24 godzin należy określić zarówno MR, jak i RQ, aby umożliwić precyzyjne obliczenie utraty/przyrostu netto konkretnego substratu (np. tłuszczów). Mimo że odżywianie, wysiłek i środowisko mogą zwiększyć BMR, RQ jest dynamiczny (nie odrębny dla każdego rodzaju wysiłku) i oddziałuje na chwilową całkowitą MR. Jeśli okresy najwyższego MR następują w czasie najwyższego RQ, magazynowane tłuszcze są chronione. Zastosowanie średniej lub przewidywanej przemiany materii w ciągu 24 godzin w porównaniu do kalorii faktycznie przyswojonych jest niewystarczające, aby dokładnie przewidzieć utratę/przyrost substratu. BMR, podstawowa przemiana materii; DIT, termogeneza indukowana pożywieniem; EE, wydatek energetyczny; MR, przemiana materii; RQ, współczynnik oddechowy
We wczesnych doświadczeniach z użyciem kalorymetru dokładnie mierzono ciepło emitowane z wyizolowanego termicznie pomieszczenia, uzyskując bezpośrednie pomiary kalorymetrem dotyczące godzin, a nawet dni.[16] Później, częściowo z przyczyn praktycznych, większość badaczy zwróciła się ku pomiarom wytworzonego dwutlenku węgla (CO2) i zużytego tlenu (O2), aby uzyskać pośrednie pomiary kalorymetrem ciepła wykształconego z procesu oddychania. Pierwsze systematyczne badanie zawartości energii brutto w żywności przypisuje się Rubnerowi w Niemczech i Atwaterowi w Stanach Zjednoczonych.[7] Porównując pośrednią kalorymetrię uczestników badania z powiększającą się bazą danych dotyczącą badań z wykorzystaniem bezpośredniej kalorymetrii bombowej i obejmującą ponad 4000 analiz 1360 różnych produktów żywnościowych, Atwater ulepszył wcześniejsze dokonania Rubnera i stworzył gamę czynników i współczynników reprezentujących dostępność energii i strawność artykułów spożywczych, aby opisać ich zawartość przydatnych kalorii.[17] Jednakże uproszczony i uogólniony pogląd o magazynowaniu energii jako konsekwencji „przyswajania i spalania kalorii”, interpretowany często jako dieta i ćwiczenia, nie ujmuje bardziej subtelnej termodynamiki magazynowania i usuwania energii, w szczególności ciepła odpadowego. Ponadto, organizm człowieka nie jest idealną maszyną i wiadomo, że energia uwolniona ze spalania żywności nie jest identyczna jak energia dostępna dla organizmu z przyswajania pożywienia.[18] Koncepcja ta, znana jako „energia metaboliczna”, oznacza różnicę między energią brutto spożytej żywności mierzonej za pomocą kalorymetrii bombowej a energią zawartą w odchodach.[19]
Aby odnieść się do dynamicznej natury przemiany materii, w niniejszej niejednorodnej, krótkiej analizie metabolizmu należy uwzględnić współczynnik oddechowy [RQ]. Można go oszacować za pomocą współczynnika wymiany oddechowej (ang. respiratory exchange ratio, RER) lub współczynnika wytworzonego CO2 i zużytego O2 podczas oddychania i można go bezpośrednio powiązać z pojedynczym substratem ulegającym utlenianiu, czy zalicza się on do białek, węglowodanów, tłuszczów czy alkoholi.[20] Ogólne obliczenia stechiometryczne dla białek, węglowodanów, tłuszczów, alkoholu i innych, mniej licznych źródeł węgla w diecie, takich jak octany i ciała ketonowe przedstawiono na Ryc. 2. Znaczenie pomiarów współczynnika wymiany oddechowej [RER] polega na możliwości wykorzystania ich do oszacowania współczynnika oddechowego [RQ] mieszaniny różnych źródeł paliwa (tj. typowego posiłku składającego się z różnego rodzaju makroelementów).[15] Ogólnie rzecz biorąc, lipidy posiadają współczynnik oddechowy [RQ] o wartości ~0,7, zaś węglowodany mają współczynnik oddechowy [RQ] o wartości ~1,0. RQ alkoholi jest nieco niższy niż lipidów i wynosi ~0,67, a podczas gdy współczynnik oddechowy [RQ] aminokwasów waha się od 0,6 do 1,17, średnia wynosi ~0,84. W przypadku białek, jako że aminokwasy nie są całkowicie utlenione, mocz i mocznik w kale zbiera się w okresie 24 godzin, aby wyliczyć utlenianie białek; wartość ta jest następnie odejmowana od pomiarów RQ w celu uzyskania RQ niebiałkowego (npRQ, ang. nonprotein RQ).[15] Przypadkowo, ponieważ wartości współczynnika oddechowego [RQ] białek ogólnie mieszczą się w środku przedziału dla węglowodanów/tłuszczów, kalorymetria pośrednia daje względnie dokładny, ilościowy obraz podziału energii z węglowodanów/tłuszczów bez równowagi azotowej[15]. Dla pełniejszych i bardziej szczegółowych ocen podziału energii, wykorzystano npRQ.
Ogólne równanie stechiometryczne utleniania
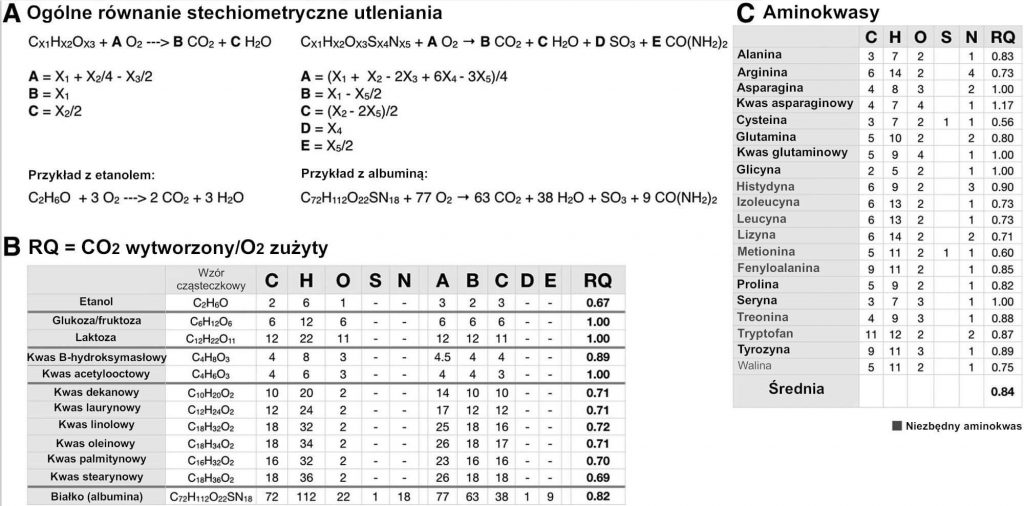
Ryc. 2.: Utlenianie i RQ substratów. (A) Ogólne obliczenia stechiometryczne dla różnych substratów służących jako paliwo. (B) RQ komórkowy – stosunek wytworzonego CO2 do zużytego O2 – dla szerokiej gamy produktów żywnościowych. (C) Podsumowane zostały poszczególne wartości RQ aminokwasów; przeciętny RQ wszystkich aminokwasów wynosi RQ = 0,84. Ogólnie przyjęte wartości współczynnika oddechowego [RQ] głównych makroelementów to białka (B) = 0,82; węglowodany (W) = 1,0; oraz tłuszcze (T) = 0,70.
Dla prawidłowej interpretacji homeostazy energii, znaczenie współczynnika oddechowego [RQ] jest nie do przecenienia. Należy wziąć pod uwagę nie tylko ilość każdego przyswojonego makroelementu, ale również, co bardziej istotne, sposób w jaki paliwa te są magazynowane i wykorzystywane. Nie można wyliczyć nadmiaru lub niedoboru tłuszczów, jeśli przeanalizuje się jedynie 24-godzinną ekstrapolację (TEE) 15-minutowego pomiaru (BMR/RMR) i porówna wynikający z tego całkowity przewidywany wydatek na aktywność fizyczną z faktyczną liczbą spalonych kalorii, o ile nie uzyska się precyzyjnej wiedzy na temat sposobu, w jaki zachodzi podział makroelementów służących jako paliwo w ciągu całego dnia.
Co istotne, współczynnik oddechowy [RQ] ujmuje wzrost i spadek procentowy spalania tłuszczów spowodowanego dietą, wysiłkiem i środowiskiem. A jeśli połączy się współczynnik oddechowy [RQ] z całkowitego dziennego wydatku energetycznego [TEE], oczywiste jest, że spalanie i magazynowanie tłuszczów są najbardziej istotne dla przyrostu lub utraty wagi; można to przedstawić za pomocą zapisu matematycznego w następujący sposób:
[ʃ(tłuszcze%)EEdt] – SPOŻYCIE TŁUSZCZÓW24 h = UTRATA/PRZYROST WAGI
Pomimo licznych badań wykazujących, że lipogeneza de novo z węglowodanów nie przyczynia się istotnie do zwiększenia się zapasów tkanki tłuszczowej w całym organizmie obecnych u osób otyłych,[21-27] rozpowszechniony jest pogląd, że „cukier zmienia się w tłuszcz”. Być może jeszcze większe nieporozumienie wynika z niezamierzonych konsekwencji łączenia węglowodanów złożonych pochodzących z pełnowartościowej żywności z wysokoprzetworzonymi zbożami i cukrami po prostu dlatego, że mają wspólne określające wiązanie glikozydowe obecne we wszystkich węglowodanach. To nie tylko odwraca uwagę od ważnych negatywnych skutków dla metabolizmu, jakie nadmierne spożycie cukrów prostych niesie dla stłuszczenia wątroby i dysfunkcji mitochondriów, ale także zaciemnia obraz roli, którą nadmiar białek (aminokwasów), alkoholi oraz węglowodanów w diecie odgrywa w doprowadzaniu do magazynowania tłuszczów w żywności i zakłóceń usuwania magazynowanej tkanki tłuszczowej poprzez normalne czynności metaboliczne na czczo.
Zatem w jaki sposób można przeformułować uogólnione poglądy dotyczące metabolizmu tak, aby zapewniały bardziej prawidłową ocenę procesu usuwania i gromadzenia tłuszczów? Rozważmy podstawową przemianę materii [BMR] na czczo i w stanie bezczynności w okresie 24 godzin, jak pokazano na Ryc. 1. W każdej godzinie postu niedobór tłuszczów się zwiększa, a jego wielkość określa odsetek tłuszczów w całkowitym wydatku energetycznym [TEE] w każdej jednostce czasu. Po przebudzeniu i przy niewyczerpanych zapasach glikogenu, normalny współczynnik oddechowy [RQ] na czczo wynosi ~0,85 lub ~50% utleniania zarówno z magazynowanych tłuszczów, jak i węglowodanów. W tym scenariuszu przewiduje się, że mniej więcej połowa spodziewanego niedoboru kalorii dla 24-godzinnego całkowitego wydatku energetycznego [TEE] pochodzi z tłuszczów. Następnie rozważmy dodatkowy wysiłek i powiązany z nim wzrost wydatku energii (EE) w okresie aktywności wraz z towarzyszącym wzrostem RQ (sprzyjającym utlenianiu węglowodanów), co jest typowe dla okresów zwiększonego wysiłku. Zauważmy, że w każdej godzinie zmienionego pod wpływem wysiłku RQ i zwiększonego wydatku energii [EE], „nowy” RQ nie tylko oddziałuje na spalane kalorie podczas dodatkowego wysiłku, ale także wpływa na kalorie, które zostałyby spalone przez podstawową przemianę materii [BMR] w tym samym okresie. Stwarza to na pozór paradoksalną sytuację, jako że możliwe jest znaczące zwiększenie wydatku energii [EE] za pomocą ćwiczeń i jednoczesne zmniejszenie wykorzystania procentowej zawartości tłuszczów do poziomu malejących zysków, kiedy porównanie dotyczy wyłącznie ograniczenia kalorii.
Załóżmy przykładowo, jak pokazano na Ryc. 3, że podstawowa przemiana materii [BMR] danej osoby wynosi 1800 kcal/dzień (75 kcal/h). Jedna godzina wysiłku fizycznego (tzn. biegania w tempie 8 km/h) przyniesie skutek w postaci wzrostu wydatku energii [EE] do ~500 kcal/h. Jednak to zużycie kalorii tłuszczowych (będące skutkiem zmian współczynnika oddechowego [RQ] w każdej dowolnej jednostce czasowej) jest bardziej znaczące i ważniejsze niż całkowity wydatek energii [EE], jeśli chodzi o wagę ciała. Jak pokazuje wiele badań z użyciem kalorymetru, dynamiczna aktywność zazwyczaj podnosi RQ i dlatego też zmniejsza procent wykorzystania tłuszczów w okresie aktywności.[9,28,29] Alternatywnie, osoba na czczo/w stanie spoczynku lub podczas długotrwałego ograniczenia kalorii będzie posiadać obniżony współczynnik oddechowy [RQ], a przez to zwiększony procent wykorzystania tłuszczów.[30,31] Pojęciowo trudne może być postawienie znaku równości pomiędzy jedną godziną biegania a trochę więcej niż połową łyżeczki oliwy z oliwek (~40 kcal/łyżeczka) w odniesieniu do ekwiwalentów energii. Jednakże spostrzeżenie to szybko wyjaśnia dlaczego wzrost wydatku energii [EE] indukowany ćwiczeniami może w krótkim czasie pokonać dieta. Chociaż często twierdzi się, że ćwiczenia zachowują masę beztłuszczową ciała, kiedy precyzyjnie połączyć ograniczenie kalorii z przyjmowaniem kalorii i ćwiczeniami, wydaje się, że utrata jest jednakowa.[11] Redman i in. sugerują ponadto, że ludzie są genetycznie lub epigenetycznie zaprogramowani do utraty i gromadzenia tłuszczów – reakcja „first on-last off”, która może wyjaśniać dlaczego obszary problematyczne, takie jak tłuszcz na brzuchu, różnią się u osób o podobnej sprawności fizycznej.[11] Finlayson i in. przypuszczają również, że korzyści z ćwiczeń mogą być zrównoważone u niektórych osób przez „nagrodę indywidualną”, polegającą na tym, że nasilające się pragnienie jedzenia równoważy wszelkie kalorie utracone podczas wysiłku.[32] Sugerujemy, że ustalanie dokładnej przyczyny zmiany wagi mogą dodatkowo maskować zmiany termodynamiczne podziału paliwa wskutek złożonej interakcji aktywności, źródła kalorii oraz środowiska.
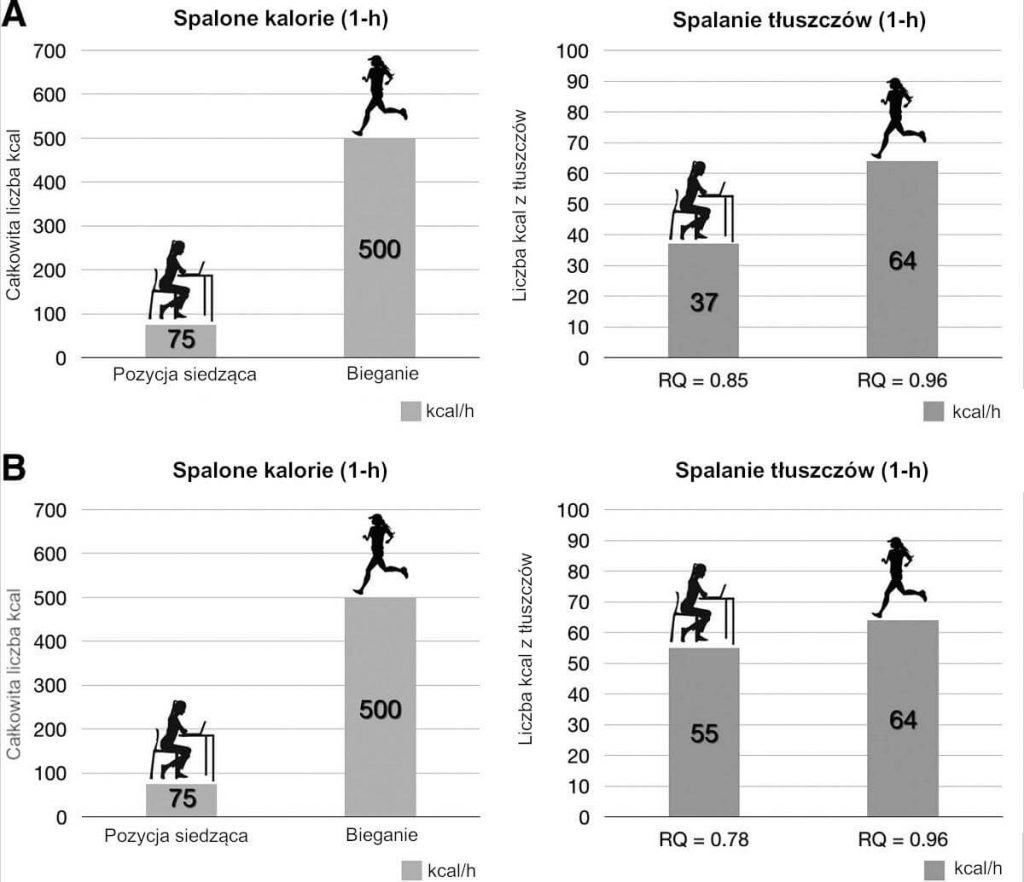
RYC. 3.: Wykorzystanie paliwa w przypadku aktywności fizycznej w przeciwieństwie do ograniczenia kalorii. Jednogodzinny blok czasowy porównano z jednogodzinnym blokiem czasowym spędzonym na bieganiu. (A) Przy przemianie materii w stanie spoczynku [RMR] wynoszącej 1800 kcal/dzień (przeciętna osoba), przewidywany współczynnik oddechowy [RQ] w stanie spoczynku, wynoszący 0,85 (49,3% utleniania tłuszczów) skutkuje wydatkiem energii [EE] o wartości 75 kcal/h i 37 kcal/h spalania tłuszczów. W czasie aktywności fizycznej wzrost RQ do 0,96 (12,8% utleniania tłuszczów) skutkuje EE o wartości 500 kcal/h i 64 kcal/h spalania tłuszczów. Dla porównania zmiana spalania tłuszczów pomiędzy stanem spoczynku a stanem aktywności równa się nieco więcej niż ½ łyżeczki oliwy z oliwek (~40 kcal/łyżeczka). (B) W okresach długotrwałego ograniczenia kalorii, RQ w stanie spoczynku może spaść do poziomu 0,78 (~74% utleniania tłuszczów) ze względu na zwiększone spalanie tłuszczów, co skutkuje spalaniem tłuszczów w wysokości ~55 kcal/h. Jest to różnica jedynie 9 kcal (mniej niż ¼ łyżeczki oliwy z oliwek) w stosunku do 64 kcal/h spalania tłuszczów przewidywanego w czasie aktywności fizycznej bez ograniczenia kalorii. EE, wydatek energii; RMR, przemiana materii w stanie spoczynku.
Termodynamika odżywiania
Odżywianie wraz z powiązaną z nim DIT i priorytetem oksydacyjnym dodaje jeszcze jeden poziom złożoności do pytania „czy kaloria jest kalorią?”. Rycina 4 ilustruje wpływ częstotliwości posiłków na czas dostępny na usunięcie przyswojonej energii. W stanie na czczo jedynym źródłem energii metabolicznej są zapasy tłuszczu. Jeżeli post trwa, zapasy glikogenu zostaną w końcu wyczerpane i nastąpi zmiana w postaci utleniania tłuszczów.33 Ketony pochodzące z kwasów tłuszczowych mogą nie tylko przyczynić się aktywności metabolicznej, ale także pokryć do 60-70% zapotrzebowania mózgu na energię w ciągu długich okresów spadku glikogenu; stężenia glukozy we krwi zachowują glukoneogeneza oraz aminokwasy glukogenne pozyskiwane z zapasów białek.[34,35]
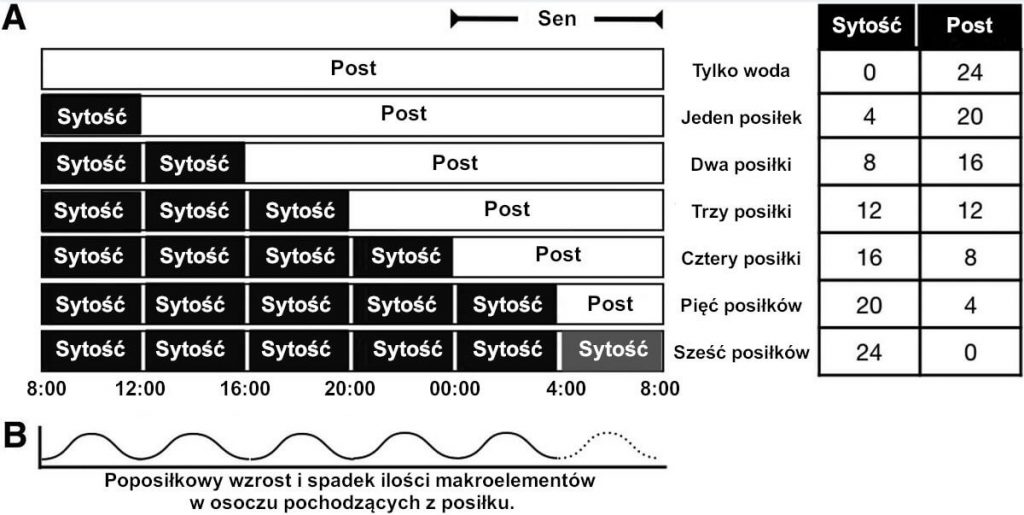
Ryc. 4.: Stan sytości i postu wobec częstotliwości posiłków. (A) Różne częstotliwości posiłków i ich wpływ na stan sytości/postu. (B) Poposiłkowe wahania poziomu makroelementów w osoczu w oparciu o częstotliwości posiłków.
Podczas jedzenia inicjowany jest szereg złożonych procesów trawiennych, które przynoszą skutek w postaci rozkładu spożytych produktów żywnościowych do ich składowych makroelementów (i mikroelementów) oraz dystrybucji tych makro- i mikroelementów do tkanek. Co istotne, wskutek uwzględnienia stanów sytości na bardziej ogólnym modelu konceptualnym, problem, który może wyniknąć w efekcie przewlekłego przejadania się staje się ewidentny. Dodatek każdego kolejnego posiłku powoduje zaburzenie stanu postu i obliguje organizm do podjęcia działania w stosunku do nowo przyswojonego paliwa. I mimo że każdy posiłek wiąże się z 4-6-godzinnym poposiłkowym zwiększeniem tempa przemiany materii[36], to, co stanie z przyswojonym paliwem jest kwestią indywidualną, jako że zostanie ono zmetabolizowane, zmagazynowane, usunięte lub poddane kombinacji tych procesów. Wszystkie przyswojone kalorie muszą ostatecznie ulec któremuś z tych trzech mechanizmów. Ponieważ posiłki składają się z mieszanki różnych makroelementów, logiczne pytanie koncentruje się na regulacji podziału paliwa po posiłku: w jaki sposób organizm decyduje które paliwo wykorzystać, usunąć lub zmagazynować?
Termogeneza indukowana pożywieniem [DIT] i priorytet oksydacyjny
Prentice i współpracownicy odnieśli się do kwestii podziału paliwa opracowując model priorytetu oksydacyjnego lub hierarchii oksydacyjnej w oparciu o indywidualne magazynowanie makroelementów (Ryc. 5).[37-44] Aby lepiej zrozumieć to, co stanie się z komponentami molekularnymi żywności, kiedy zaczną opuszczać przewód pokarmowy, konceptualnie użyteczne może być odłożenie na bok powszechnego nazywania żywności i schematu porządkowania makroelementów takich jak białka, węglowodany i tłuszcze oraz uznanie, że całe produkty żywnościowe, zwłaszcza posiłki jako całość, są mieszankami. Węglowodany złożone i cukry proste rozkładane są do monosacharydów, białka do aminokwasów składowych, a lipidy mobilizowane na drodze transportu przez chylomikrony. Tak więc jedzenie skutkuje poposiłkowym wzrostem poziomu monosacharydów (glukozy), aminokwasów oraz lipidów we krwi, podczas którego organizm stara się znormalizować ich stężenia za pomocą jednej z trzech strategii: wykorzystania, magazynowania lub usuwania. Co ważne, gdy magazynowanie jest ograniczone lub niemożliwe, pozostają dwie opcje: wykorzystanie bądź usuwanie.
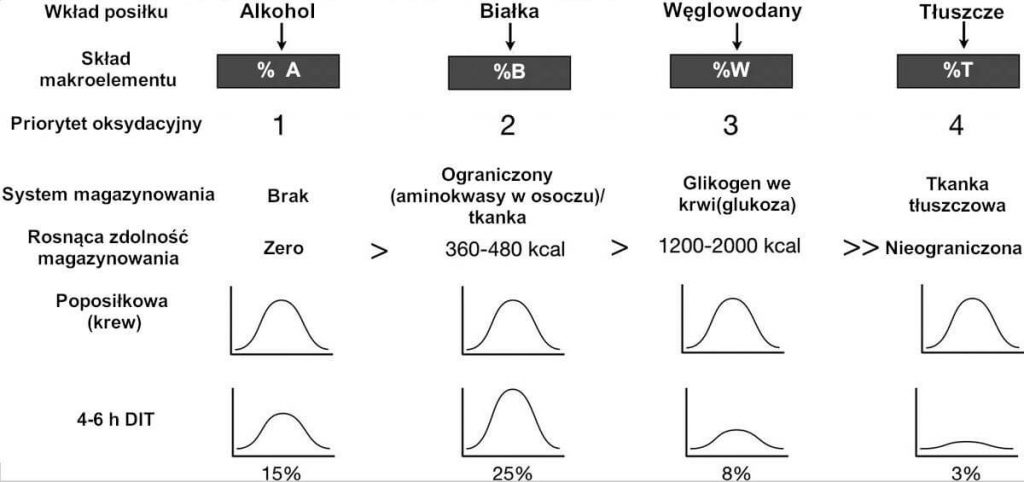
RYC. 5.: Priorytet oksydacyjny usuwania makroelementów. Priorytetyzacja oksydacyjna makroelementów przewiduje hierarchię podziału opartą na odwrotnej relacji zdolności magazynowania. Nie wszystkie makroelementy mogą być magazynowane na tym samym poziomie. Dla przykładu alkohole nie posiadają długofalowej zdolności magazynowania, podczas gdy zdolność tłuszczów jest relatywnie nieograniczona. Chociaż wzrost DIT po posiłku jest największy dla białek i najmniejszy dla tłuszczów, ów wzrost metabolizmu niekoniecznie odzwierciedla spalanie magazynowanych rezerw energii i lepiej wyjaśnia je potrzeba minimalizowania poposiłkowego wzrostu poziomów substancji odżywczych we krwi. Niemal we wszystkich przypadkach poposiłkowe poziomy analitów we krwi, które rosną na skutek spożycia posiłku normalizują się w ciągu 4-6 godz., kiedy osiągany jest stan postabsorbcyjny. A, alkohol; W, węglowodany; DIT, termogeneza indukowana pożywieniem; T, tłuszcze; B, białka.
Przykładowo po spożyciu posiłku bogatego w białka obserwuje się relatywnie większy wzrost termogenezy indukowanej pożywieniem [DIT] w stanie po posiłku w porównaniu z posiłkiem bogatym w węglowodany, jako że aminokwasy nie posiadają istotnego dziedzicznego magazynu; organizm w zasadzie przetwarza nadmierną energię w ciepło odpadowe. Mimo że poposiłkowy wzrost metabolizmu po spożyciu posiłku bogatego w białka można uznać za korzystny i potraktować jako sposób złagodzenia przyrostu wagi czy nawet sposób na jej utratę, niewielka ilość tej nadmiernej energii, jeśli w ogóle jakakolwiek, pozyskiwana jest z magazynowanej tkanki tłuszczowej. Jak pokazuje wzrost tempa przemiany materii na skutek aktywności fizycznej, możliwe jest, a nawet spodziewane, że szybszy metabolizm w celu usunięcia nadmiaru kalorii przyswojonych z posiłku bogatego w białka może faktycznie spowodować mniejsze spalanie magazynowanej tkanki tłuszczowej. Równie istotny jest następujący czynnik: nie tylko tłuszcze z pożywienia mogą być magazynowane w szybszym tempie w obecności makroelementów o wyższym priorytecie oksydacyjnym, ale także ogólne współczynniki utleniania tłuszczów mogą spowolnić zmiany w podziale paliwa.[45]
W przeciwieństwie do tego silniejsza odpowiedź termogeniczna na pożywienie nie zawsze musi powodować, że rezerwy nadmiaru magazynowanej energii zostaną spalone. Koncepcję tę udowadniają dane pokazujące skutki podania glukozy i etanolu bezpośrednio w zastrzyku dożylnym.[45] Podczas gdy zastrzyki etanolu i glukozy nie mają znaczącego wpływu na ogólną przemianę materii w stanie spoczynku [RMR], sam etanol obniża utlenianie tłuszczów o 87%. Co istotne, zastrzyk glukozy po zastrzyku etanolu przewidywalnie nasila utlenianie węglowodanów o 249% i jeszcze bardziej obniża utlenianie tłuszczów niemal do zera przez 90 min., przy czym później zwiększanie utlenienia tłuszczów przebiega powoli; w rzeczywistości całkowite 4-godzinne zintegrowane utlenianie tłuszczów w następstwie zastrzyku etanolu, a następnie glukozy jest obniżone o 79%.
Wracając do kwestii częstotliwości posiłków i priorytetu oksydacyjnego różnorodnych posiłków, wyniki te można odnieść do tego, co dzieje się, kiedy osoba spożywa przekąskę w postaci wina, sera i krakersów. Wzrost stężenia alkoholi i glukozy po spożyciu przekąski faktycznie obniża utlenianie tłuszczów i sprzyja magazynowaniu tłuszczów z pożywienia. Skoro tak, łatwo zrozumieć w jaki sposób tkanka tłuszczowa w organizmie zwiększa się wraz z upływem czasu; presja homeostatyczna do znormalizowania wzrostu poziomów składników odżywczych we krwi po posiłku zapewnia nowatorski, konceptualny model, który przewiduje akumulację tłuszczów.
Termogeneza adaptacyjna
Dodatkowym elementem, który wpływa na metabolizm organizmu jest ogólnie postrzegane środowisko – w szczególności łagodny stres zimna.[46-49] W przeciwieństwie do aktywności fizycznej, która zwiększa współczynnik oddechowy [RQ], środowisko, w jakim znajduje się organizm może indukować przeciwny efekt.[29,50] Przykładowo długie okresy łagodnego stresu zimna wynikające z niższej temperatury otoczenia mogą indukować termogenezę bezdrżeniową, zmniejszając RQ (sprzyjający utlenianiu tłuszczów)51; zwiększa to jednocześnie wydatek energii [EE] w stanie postu/w stanie spoczynku i może prowadzić do ogólnego, zwiększonego deficytu tłuszczów.
W szczególności aktywacja brązowej [brunatnej] tkanki tłuszczowej (ang. BAT, brown adipose tissue) zwiększa produkcję ciepła przez mitochondria w komórkach mięśni szkieletowych,[50,52-54] przynajmniej częściowo poprzez regulację pozytywną receptora aktywowanego przez proliferatory peroksysomów, koaktywator gamma 1-alga (ang. PGC-1α), który indukuje niską podaż kalorii lub łagodny stres zimna.[52,53,55-58] Kwestię dlaczego PGC-1α ulega regulacji pozytywnej w odpowiedzi zarówno na niską podaż kalorii trzeba jeszcze w pełni wyjaśnić; jednak w naszej „Metabolic Winter Hypothesis”1 postulujemy, aby odpowiedzi cząsteczkowe na te dwa stresory (tj. ograniczenie kalorii oraz niskie temperatury) mogły wykształcić się w tandemie – obie konserwują energię w stanie postu i transportują kalorie do komórek tłuszczowych, by uzyskać ciepło w okresach zimna. Co więcej, zgodnie z tym, co zgłaszają zarówno Valdés i in.[59], jak i Yang i in.[60], którzy badali względnie jednorodne populacje genetyczne, spożywające porównywalne pożywienie, żyjące na umiarkowanie różnych szerokościach geograficznych, istnieje związek między temperaturą otoczenia a otyłością.
Ogólnie rzecz biorąc, dyskusja na temat „przyswajania i spalania kalorii” niejednokrotnie uwzględnia uogólnioną przemianę materii, termogenezę indukowaną pożywieniem [DIT], energię związaną z aktywnością fizyczną, a nawet brązową tkanką tłuszczową [BAT] i łagodnym stresem zimna. Jednakże, koniec końców, to zintegrowana energia całkowita i podzielone paliwo (przemiana materii i RQ) określają ile tkanki tłuszczowej zużywa w danym okresie 24 godzin. W okresach postu z wyczerpaniem związków innych niż glikogen, podział węglowodany:tłuszcze (RQ~0,85) może wynosić 50:50. W stanie po posiłku/sytości wykorzystanie tłuszczów może być istotnie zredukowane; w przeciwnym razie, w okresie przeciągającego się postu (ketoza z wyczerpaniem glikogenu), wykorzystanie tłuszczów może dominować. Oczywiście zwyczajne, przeciętne zestawienie danych dotyczących przemiany materii oraz wszechobecny nacisk na diety i ćwiczenia „zwiększające” metabolizm nie wystarczają, by wyjaśnić termodynamikę żywienia; co ważne, mogą przynieść niezamierzony skutek w postaci modyfikacji przemiany materii i nieoczekiwanych zmian RQ nie sprzyjających spalaniu tłuszczów.
Piramida żywności i plany dietetyczne uwzględniające pełnowartościowe pożywienie
Jak już wcześniej wyjaśniliśmy bardziej szczegółowo, [1] piramida żywności stanowi nowatorską metodę konceptualizacji pożywienia w sposób, który eliminuje tradycyjne, będące w powszechnym użyciu grupy makroelementów żywności, czyli białek, węglowodanów i tłuszczów. Piramida żywności organizuje pełnowartościowe pożywienie z wykorzystaniem paradygmatu gęstości energii. Uznaje, że niezbędne aminokwasy (tj. białka) nie są ograniczającymi składnikami odżywczymi w żadnej diecie pełnowartościowej, która spełnia dzienne zapotrzebowanie na energię. Właściwie aminokwasy pochodzące z warzyw lub pochodzenia zwierzęcego, które przekraczają dzienne zapotrzebowanie, wraz z węglowodanami i tłuszczami stają się paliwem, przy czym priorytet oksydacyjny nadawany jest temu makroelementowi, który nie jest łatwo magazynowany. Taka organizacja gęstości energii pozwala poszczególnym osobom odnieść się do swoich wymogów dotyczących mikroelementów (wierzchołek piramidy), nie doprowadzając do przewlekłego przejadania się (dolne rogi piramidy). Te produkty żywnościowe stają się żywieniową podstawą codziennych posiłków, nie zaś alternatywą o większej gęstości energii. Zapewniają również bogate źródło fitoskładników i mogą być spożywane niemal bez ograniczeń. Piramida ponadto kładzie nacisk na produkty, które mają coraz większe znaczenie dla zdrowia mikrobioty jelitowej.
Wykorzystując piramidę żywności oraz łącząc efekt okresów sytości/postu, zmian RQ indukowanych pożywieniem i aktywnością fizyczną oraz priorytetu oksydacyjnego, możliwe jest przewidzenie spożytkowania substratów i tego jak różne plany dietetyczne skutecznie wpływają na preferencyjne usuwanie tłuszczów (Ryc. 6). Chociaż nad znaczeniem wpływu, jaki na przyrost masy tkanki tłuszczowej ma spożywania węglowodanów w porównaniu do spożywania tłuszczów deliberuje się od dziesięcioleci, piramida żywności zapewnia ramy konceptualne związku między dietą a gęstością energii i sugeruje, że jedno i drugie (tj. węglowodany [cukry] i tłuszcze w pożywieniu) z osobna i synergistycznie przyczynia się do przyrostu masy tkanki tłuszczowej. Uznajemy, że złożone, współzależne role insuliny, glukagonu oraz inkretyn[61,62] – nie zapominając o innych hormonach i cząsteczkach regulatorowych – zarówno w stanie postu, jak i po posiłku zapewniają mechaniczny wgląd w sposób, w jaki priorytet oksydacyjny jest zarządzany, przy czym nie zostały one kompleksowo omówione w niniejszym opracowaniu.
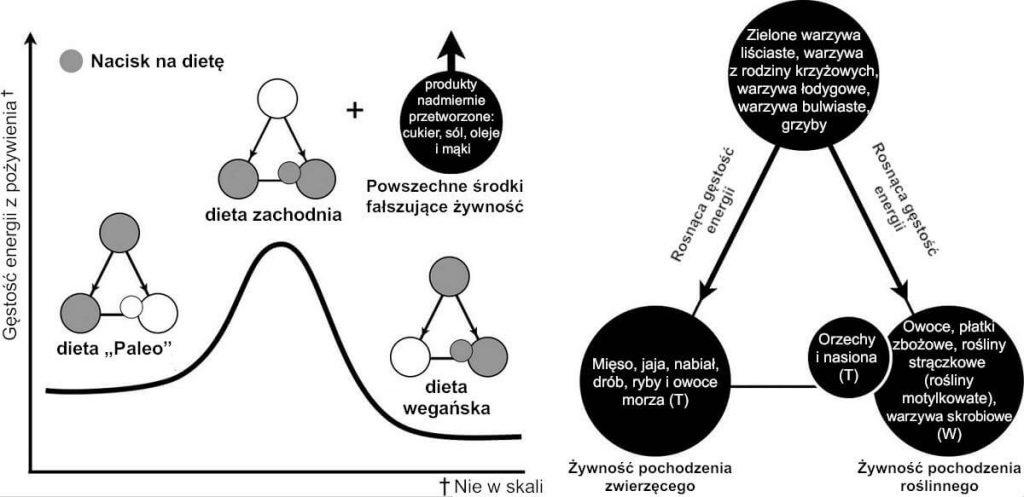
Ryc. 6: Gęstość energii popularnych programów dietetycznych. Nasza wersja piramidy żywności może być wykorzystana do porównania gęstości energii popularnych programów dietetycznych pod względem gęstości energii. Dla przykładu pokazano relatywną gęstość energii diety Paleo w porównaniu z dietą wegańską, które niemal odzwierciedlają przeciwne strony piramidy żywności. Co więcej, przewiduje się, że „dieta zachodnia”, która bardziej przemawia na korzyść produktów żywnościowych znajdujących się u podstawy piramidy żywności skutkuje maksymalnym gromadzeniem energii tłuszczów z pożywienia. Choć nie są uznawane za pełnowartościowe produkty włączonych do piramidy żywności, nadmiernie przetworzone olej, cukier i środki fałszujące żywność dodawane do zbóż (np. do mąki) mogą sprawić, że każdy z tych programów dietetycznych doprowadzi do przewlekłego przejedzenia. Podczas gdy każda strona piramidy żywności umożliwia przewidzenie nadmiaru energii, można ją także wykorzystać do prognozowania nadmiaru aminokwasów i tłuszczów oraz niedoboru fitoskładników i błonnika z pożywienia.
Jednakże naszym celem w niniejszym opracowaniu nie jest przedstawienie szczegółowego omówienia biochemii i endokrynologii żywieniowej; jest nim raczej (1) skupienie się na kwestii dlaczego wpływ społeczny i nienaturalny dostęp do pożywienia stworzyły okazję do chronicznego stanu po posiłku i (2) organizacja pożywienia w zrozumiały sposób, który umożliwia dużo łatwiejsze przewidzenie gromadzenia tłuszczów. Te pojęciowe ramy wyjaśniają również dlaczego diety, które eliminują jeden bądź drugi makroelement (tj. diety, które eliminują tłuszcze, ale uwzględniają węglowodany lub vice versa) mogą być skuteczne na krótką metę, lecz zazwyczaj nie przynoszą skutku w kontekście długotrwałej utraty wagi, kiedy wyeliminowany produkt żywnościowy zostaje przywrócony. Wierzchołek piramidy żywności zawiera fitoskładniki/bogate w błonnik produkty niskokaloryczne. Wzdłuż każdego opadającego boku piramidy żywności znajdują się produkty o rosnącej gęstości energii, podzielone głównie na produkty będące źródłem tłuszczów i węglowodanów. Godnym uwagi wyjątkiem są tłuszcze pochodzące z pełnowartościowych produktów roślinnych, takich jak orzechy, nasiona oraz konkretne owoce (np. awokado i orzechy kokosowe).
Jeżeli spożywamy produkty z właściwej strony piramidy żywności, pozyskując energię przede wszystkim z pełnowartościowej skrobi pokarmowej i owoców, węglowodany pokarmowe (oprócz tych potrzebnych do uzupełnienia zapasów glikogenu) mogą być usunięte poprzez zwiększony współczynnik oddechowy [RQ] i wydatek energii [EE] po posiłku przez termogenezę indukowaną pożywieniem [DIT]. Dieta pochodząca z lewego boku piramidy żywności, przeciwnie, prowadzić będzie do zubożenia zapasów glikogenu i ostatecznie spowoduje stan ketozy, doprowadzając do spadku RQ i wspierając wykorzystanie tłuszczów. W odróżnieniu od nadmiaru węglowodanów pokarmowych, które usuwane są na drodze utleniania, nadmiar lipidów pokarmowych może być częściowo eliminowany w kale po niepełnym wchłonięciu. Każde podejście może być skuteczne, jeśli chodzi o spowodowanie niedoboru kalorii wymaganego do osiągnięcia efektu, jakim jest utrata wagi; jednakże długofalowe decyzje dotyczące stylu życia, aby jak najdłużej zachować zdrowie są bardziej złożone. A zatem piramida żywności zasadniczo zapewnia model zdrowego odżywiania, który wykracza poza skład kaloryczny artykułów spożywczych.
Diety, które kładą nacisk na podstawę piramidy żywności – być może nawet jako niezamierzony skutek tego, co można nazwać „zbilansowanym posiłkiem” – mogą doprowadzić do nadmiernego magazynowania tłuszczów z wykorzystaniem mechanizmów priorytetu oksydacyjnego opisanych powyżej. I, jak pokazano na Ryc. 6, model priorytetu oksydacyjnego prognozuje, że poposiłkowy podział i usuwanie artykułów spożywczych w typowej diecie „zachodniej” (tj. diecie kładącej nacisk na podstawę piramidy żywności) będą powodować tycie; owa prognoza odpowiada aktualnej rzeczywistości.[63] Spożywanie produktów znajdujących się na dole piramidy żywności, zwłaszcza w przypadku dodatkowej konsumpcji przetworzonych ziaren (np. różnego rodzaju mąki) i środków fałszujących olej i cukier, daje dietę, w której większość produktów żywnościowych o największej gęstości energii konsumowanych jest łącznie. Wiele tradycyjnych planów posiłków sprzed wieku lub wcześniejszych miało na celu zminimalizowanie ekonomicznego niedoboru kalorii, ale mogły one w sposób niezamierzony doprowadzić do obecnego szeroko rozpowszechnionego, indukowanego kulturowo odżywiania się produktami znajdującymi się na dole piramidy żywności (np. burger-frytki, stek-ziemniaki, ryba-frytki, makaron-sos mięsny, curry-ryż itd.). Jednak w środowisku nienaturalnej dostępności kalorii, posiłki te stają się źle przystosowane.
W połączeniu z możliwym do przewidzenia priorytetem oksydacyjnym i regulowaną zdolnością do magazynowania nadmiaru węglowodanów jako glikogenu sprzyja to magazynowaniu tłuszczów do późniejszego użytku; niestety dla większości późniejszy użytek nigdy nie nadchodzi. Współczesne sukcesy w rolnictwie, zmniejszony koszt żywności i dostępność – w połączeniu z aktualnymi modelami snu – umożliwiają stan przewlekłej sytości; zaś biorąc pod uwagę nadmierną częstotliwość posiłków/przekąsek, możliwe jest, że wiele osób pozostaje w stanie poposiłkowym/absorpcyjnym przez większość dnia. Jest to okres, w którym priorytet ma usuwanie przyswojonych kalorii, nie zaś spalanie zapasów energii.
Stan chronicznej sytości jeszcze bardziej wspiera powszechny pogląd, że „odżywianie to stan wyjątkowy” i że częste jedzenie jest korzystne, pomimo licznych badań wskazujących na zbawienne efekty okresowego postu, diet, które naśladują post, ograniczenia kalorii, ograniczenia białka oraz długotrwałych postów wodnych pod nadzorem lekarza.[64] Jednak ograniczenie kalorii, post co drugi dzień i post okresowy mają nie tylko długą historię w społeczeństwie, ale są też jedynymi mechanizmami, za pomocą których życie/zdrowie uległy wydłużeniu w laboratorium.[65-70]
Poza tym zarówno badacze, jak i dietetycy w dalszym ciągu koncentrują się na dyskusji „jeść mięso czy nie jeść mięsa” lub „tłuszcze czy cukry”. My sugerujemy, że stanowi to fałszywy, ideologiczny rozdźwięk, który jedynie luźno odnosi się do gromadzenia tłuszczów wraz z upływem czasu. Dla przykładu gdyby ktoś miał stosować rygorystyczną dietę wegańską w oparciu o prawą stronę piramidy żywności, tym samym eliminując wszystkie tłuszcze pokarmowe pochodzące z produktów zwierzęcych, ale nie ograniczając olejów, przetworzonych ziaren (np. różnego rodzaju mąki) albo cukrów, wciąż możliwe byłoby magazynowanie tłuszczów pokarmowych. Podobnie diety z lewego boku piramidy żywności mogą powodować magazynowanie tłuszczów pokarmowych mimo często przytaczanych korzyści zdrowotnych, jakie daje utrzymywanie stanu relatywnej ketozy. Dlaczego tak jest? Ponieważ nie potrzeba dużej ilości węglowodanów pokarmowych, nawet ze zdrowych źródeł, takich jak owoce i pełnowartościowa, niskoglikemiczna skrobia, ażeby wyeliminować ketozę i zwiększyć poposiłkowy współczynnik oddechowy [RQ] i przez to wesprzeć usuwanie węglowodanów. W tym przypadku, pomimo że węglowodany pokarmowe są zwykle postrzegane jako winowajcy magazynowania tłuszczów, w rzeczywistości magazynowany jest składnik tłuszczowy posiłku. Hall i in. zgłosili również, że stosowaniu diety izokalorycznej o niskiej zawartości węglowodanów w porównaniu z wysokowęglowodanową dietą podstawową nie towarzyszy szybsza utrata tkanki tłuszczowej pomimo względnie niewielkiego wzrostu wydatku energii [EE] lub też przewagi metabolicznej.[71]
Tworzy to dietetyczną sytuację patową i chociaż składniki energii są tylko jednym aspektem piramidy żywności, w jasny sposób prognozują akumulację nadmiaru tkanki tłuszczowej, kiedy połączy się je z modelem pojęciowym priorytetu oksydacyjnego i godzinami spędzonymi w stanie sytości w przeciwieństwie do stanu postu. Dwa główne paliwa pokarmowe, tłuszcze i węglowodany, wykorzystują w zasadzie rywalizujące ze sobą próby sił związane z nadmiarem energii: kalorie z węglowodanów pokarmowych eliminują energię z magazynowanych tłuszczów, która zostałaby spalona w stanie postu i tłuszcze pokarmowe gromadzą się, zwiększając ciężar tłuszczów, który musi być wykorzystany w przyszłości. Oczywiście w stanie przewlekłej sytości, te dwa zachwiania równowagi energii sprzyjają akumulacji netto tkanki tłuszczowej, a w świecie z ograniczoną liczbą kalorii byłaby to adaptacyjna przewaga ewolucyjna dla nieczęstych okazji, by coś zjeść. Jednakże większość mieszkańców krajów rozwiniętych nie żyje w świecie z ograniczoną liczbą kalorii. Kolejną korzyścią, którą daje skierowanie dyskusji z powrotem na pełnowartościowe pożywienie z wykorzystaniem piramidy żywności zamiast dzielić składniki diety na zbyt uogólnione kategorie, takie jak białka, węglowodany i tłuszcze jest to, że potrzeba więcej szczegółów w przypadku każdego z tych makroelementów (np. niezbędne aminokwasy w przeciwieństwie do nie niezbędnych, monosacharydy proste w przeciwieństwie do węglowodanów złożonych i błonnika pokarmowego oraz kwasy tłuszczowe nasycone i trans-nienasycone w przeciwieństwie do jedno- i wielonienasyconych kwasów tłuszczowych), aby zbadać i ocenić ich wpływ na zdrowie.
Następstwa dla chorób kardiometabolicznych
Coraz częstsze występowanie nadwagi i otyłości przez ostatnich kilka dekad jest porównywalne do coraz częstszego występowania chorób kardiometabolicznych.[69,7273] Chociaż korelacja między nadwagą a chorobami kardiometabolicznymi nie jest wystarczająca, by wykazać związek przyczynowy, większość badaczy zgadza się, że te dwie kwestie są ze sobą ściśle powiązane. Co więcej, utrata wagi w wyniku działań dotyczących stylu życia, takich jak dieta i ćwiczenia, pozostaje kamieniem milowym dla terapii chorób kardiometabolicznych. Jednakże relatywny wpływ diety i ćwiczeń na utratę wagi pozostaje kwestią sporną.[74,75]
Ponadto, chociaż potencjalne zmiany nawyków żywieniowych i współczynnik oddechowy [RQ] indukowane ćwiczeniami mogą prowadzić do mniejszego spalania tłuszczów w krótkiej perspektywie czasu, nie powinny przyćmiewać niezliczonych korzyści zdrowotnych, jakie dają ćwiczenia ani zniechęcać do aktywności fizycznej. Jednak możliwość odłożenia na później lub zawieszenia nadmiernej aktywości fizycznej w okresie zmniejszonej dostępności składników pokarmowych (np. w okresie ograniczenia kalorii pokarmowych) jest realną alternatywą dla zwiększenia podaży składników pokarmowych, aby zaspokoić zapotrzebowanie metaboliczne podczas ćwiczeń. Ograniczone odżywianie w połączeniu ze zwiększoną aktywnością fizyczną może w sposób niezamierzony spowolnić powrót do kondycji i w zależności od postury danej osoby, nadmiar tkanki tłuszczowej może zwiększyć ryzyko kontuzji. Jednakże problemy te ulegają złagodzeniu, kiedy dana osoba zbliża się do normalnej wagi.
Z perspektywy ram pojęciowych istotne jest także rozpoznanie dwóch ważnych kwestii: (1) chociaż zarówno ograniczenie kalorii, jak i ćwiczenia mogą prowadzić do utraty wagi, adaptacja i reakcje fizjologiczne na jedno i drugie drastycznie się różnią i (2) niezależne od utraty wagi skutki ograniczenia kalorii i ćwiczeń dla czynników ryzyka choroby sercowo-naczyniowej nie są addytywne.76 Niemniej, ponieważ brak aktywności fizycznej i niska wydolność tlenowa są niezależnymi czynnikami ryzyka choroby sercowo-naczyniowej, ćwiczenia – o ile wykonywane są z zachowaniem bezpieczeństwa dla odpowiedniej wagi – mogą przynieść korzyść, której nie da się osiągnąć jedynie przez ograniczenie kalorii.[76]
Jak zweryfikowano powyżej, reakcje termodynamiczne na spożyte artykuły spożywcze wyraźnie różnią się od aktywności fizycznej. Pomimo że wiele osób uważa ćwiczenia za antidotum na nadmierny dopływ kalorii oraz że aktywność fizyczna bez zmian diety może prowadzić do istotnej utraty wagi, dostępne dane sugerują coś innego.[11,77-79]. Ogólnie mówiąc, negatywny wpływ współczesnych diet na wagę netto przewyższa wpływ pozytywny. I mimo że czynniki, które prowadzą do otyłości i elementy zespołu metabolicznego, które często (ale niekoniecznie) towarzyszą otyłości nie są identyczne, i tak wiążą się one z ryzykiem choroby kardiometabolicznej.[80-82]
Zakończenie
Z ewolucyjnej perspektywy adaptacyjnej wydaje się mało prawdopodobne, że nasi przodkowie mieli aż tak łatwy dostęp do społecznie normalnej i nadmiernej w obecnych czasach częstotliwości posiłków oraz gęstości kalorii. Chociaż okresy postu trwające dni lub być może nawet tygodnie mogą się obecnie wydawać społecznie ekstremalne, są możliwe do zniesienia[83-88] i były prawdopodobnie niezbędne do przetrwania w środowisku, nad którym historycznie ludzie mieli niewielką kontrolę lub nie posiadali jej wcale. Co więcej, okresowi i całodobowi dawcy czasu powodują uruchomienie silnych mechanizmów sprzężenia zwrotnego na poziomie funkcji komórkowych i ogólnoustrojowych, łącznie z wpływem na usuwanie składników pokarmowych.[89-95] Nasza „hipoteza zimy metabolicznej” sugeruje, że zdrowy tryb życia wymaga od większości z nas, abyśmy zrewidowali nasze nawyki związane z odżywianiem, snem i temperaturą naszego otoczenia.1 Wiele obserwacji wskazuje, że ograniczenia diety [DR], łagodny stres zimna oraz sen wydają się pełnić stosowną mechaniczną rolę w poprawie stanu zdrowia, wydłużeniu życia oraz minimalizowaniu chorób związanych z wiekiem.[96,97] Można to konceptualnie sformułować przeciwstawiając metabolicznej zimie (tj. chłodny, ciemny, nieruchomy i niedostateczy) metaboliczne lato (tj. ciepły, jasny, aktywny i dostateczny). Do tej struktury dochodzą niezliczone komórkowe szlaki sygnalizacyjne i podstawowe procesy komórkowe modyfikowane przez dostępność składników pokarmowych i sygnalizację tych składników. Stanowi to prawdopodobnie część szerszej fenologii mającej wpływ na całokształt życia zwierząt i roślin – zjawisko, które na skutek działań człowieka zostało skutecznie wyeliminowane z naszego codziennego życia.
Co istotne, wykorzystanie piramidy żywności w miejsce tradycyjnej kategoryzacji produktów żywnościowych jako białek, węglowodanów czy tłuszczów pozwala na łatwiejsze zidentyfikowanie wzorców żywienia, które prawdopodobnie prowadzą do przyrostu wagi. Ponadto, artykuły spożywcze po prawej stronie piramidy żywności zawierają składniki, które według badań nad długością życia mogą zapewnić zdrowie na dłużej, jeśli nie przedłużyć życie ssaków i potencjalnie ludzi98 przez minimalizowanie niezbędnych aminokwasów, co służy obniżaniu sygnalizacji szlaku mTor i GH/IGF-I, a jednocześnie zwiększanie do maksimum ilości błonnika pokarmowego, witamin, minerałów i mikroelementów, takich jak karotenoidy, fitosterole i flawonoidy, by aktywować sirtuiny i AMPK.[99-101]
Jeżeli dostępność składników pokarmowych odgrywa kluczową rolę w okresowej sygnalizacji środowiskowej, może pojawić się pytanie w jaki sposób rozmaite formy ograniczenia diety [DR] wpływają na ogólny podział i usuwanie paliwa. Przykładowo ograniczenie metioniny – obecnej w wysokich stężeniach w dietach uwzględniających białka pochodzenia zwierzęcego (lewa strona piramidy żywności) i w niskich stężeniach w dietach zawierających białka pochodzenia roślinnego (prawa strona piramidy żywności) – kojarzone jest z dłuższym życiem i lepszym zdrowiem w starszym wieku.102-104 Można również zapytać czy istnieje różnica w metabolicznym sprzężeniu zwrotnym pomiędzy pełnowartościową, bardzo niskokaloryczną dietą (ang. VLCD, very low-calorie diet), opartą na produktach roślinnych, w której ograniczone zostały substancje odżywcze, a często obficie uzupełnianymi, nowoczesnymi VLCDs (np. dietami płynnymi). Co więcej, nie jest jasne z punktu widzenia adaptacyjnej presji ewolucyjnej w jaki sposób jakikolwiek przodek człowieka mógł doświadczyć okresów poważnego niedoboru kalorii, a jednak mieć dostęp do wystarczającej ilości „uzupełniających” składników pokarmowych.
Na zakończenie słowa, które Carl von Voit napisał ponad stulecie temu w hołdzie żałobnym po śmierci Maxa Pettenkofera[7]:
Wyobraźcie sobie nasze zmysły jako obraz niezwykłych procesów metabolizmu odsłaniających się przed naszymi oczami, a przy tym poznajemy mnóstwo nowych faktów! Ustaliliśmy, że w stanie głodówki spalane są jedynie białka i tłuszcze, że podczas pracy spalane jest więcej tłuszczów i że mniejsza ilość tłuszczów spalana jest w czasie odpoczynku, zwłaszcza we śnie; że mięsożerny pies może przeżyć na diecie składającej się wyłącznie z białka, a kiedy do takiej diety białkowej doda się tłuszcze, zostaną one niemal w całości odłożone w organizmie; że węglowodany, przeciwnie, ulegają spaleniu niezależnie od ilości i że podobnie jak tłuszcze w pożywieniu, chronią organizm przed utratą tłuszczów, chociaż dopływ węglowodanów musi być większy niż tłuszczów, aby osiągnąć ten cel; że metabolizm w organizmie nie jest proporcjonalny do podatności substancji na spalanie poza organizmem, ale najłatwiej metabolizuje się białka, które spalają się z trudem na zewnątrz, a następnie węglowodany, podczas gdy tłuszcze, które z łatwością spalają się na zewnątrz, najtrudniej spalają się w organizmie.
Metabolizm jest w rzeczy samej dynamicznym procesem podlegającym wielu regulatorom, które determinują częściowo to, co stanie się z przyswojonymi kaloriami. Nasza wersja piramidy żywności przewiduje priorytet oksydacyjny spożytych produktów żywnościowych i przedstawia model pojęciowy zdrowego odżywiania wspierany badaniami dotyczącymi zdrowia i długości życia. Co ważne, na termodynamikę żywienia wpływ wywiera także stan sytości i postu, zaś zmiany współczynnika oddechowego [RQ] indukowane ćwiczeniami w zróżnicowany sposób oddziałują na usuwanie składników pokarmowych. Typowy, współczesny stan przewlekłej sytości/poposiłkowy w połączeniu z priorytetem oksydacyjnym i obfitością niedrogich produktów żywnościowych o dużej gęstości kalorii oraz awersja do łagodnego stresu zimna łączą się, by przewidywać, o ile się utrzymają się aktualne trendy, przyrost wagi w przypadku większości rozwiniętego świata.
Zobacz na: Przełom w dziedzinie tkanki tłuszczowej – dr Ellington Darden
Zasada wielkości Hennemana – zastosowanie w rozbudowie masy mięśniowej
Hipertrofia mięśniowa – Oparte na dowodach rekomendacje dotyczące treningu oporowego
Platforma szachowa mierzy kalorie spalone podczas gry w szachy online
Słowa Alergia i Anafilaksja zostały utworzone, aby opisać szkody po szczepionkach
Ludzie zaczynają się zmieniać w czterech różnych sytuacjach
Ludzie zmieniają się, gdy są wystarczająco zranieni.
Kiedy widzą wystarczająco dużo, że są do tego zainspirowani.
Kiedy nauczą się wystarczająco dużo, że chcą, i kiedy otrzymają wystarczająco dużo, że są w stanie.
To są cztery okresy, w których ludzie się zmieniają.
Bibliografia:
1.Cronise RJ, Sinclair DA, Bremer AA. The “metabolic winter” hypothesis: A cause of the current epidemics of obesity and cardiometabolic disease. Metab Syndr Relat Disord 2014;12:355–361
2.Atwater W. Discussion of the terms digestibility, availability, and fuel value. 12th Annual Report. Storrs. Agricultural Experimental Station. Storrs, Connecticut. 1900:69
3.Secor SM. Specific dynamic action: A review of the postprandial metabolic response. J Comp Physiol B 2009;179:1–56
4.Hall KD, Heymsfield SB, Kemnitz JW, et al. Energy balance and its components: Implications for body weight regulation. Am J Clin Nutr 2012;95:989–994
5.Rubner M. The Laws of Energy Consumption in Nutrition: DTIC Document; 1968
6.Benedict FG, Carpenter TM. Food Ingestion and Energy Transformations: With Special Reference to the Stimulating Effect of Nutrients. Washington, DC: Carnegie Institution of Washington; 1918
7.Lusk G. The Elements of the Science of Nutrition. Phliadelphia and London: WB Saunders Company; 1909
8.DuBois E. Basal Metabolism in Health and Disease. Philadelphia: Lea and Febiger; 1936
9.Benedict FG, Cathcart EP. Muscular Work: A Metabolic Study with Special Reference to the Efficiency of the Human Body as a Machine Washington, DC: Carnegie Institution of Washington; 1913
10.Benedict FG, Miles WR, Roth P, et al. Human Vitality and Efficiency Under Prolonged Restricted Diet. Vol. 280 Washington, DC: Carnegie Institution of Washington; 1919
11.Redman LM, Heilbronn LK, Martin CK, et al. Effect of calorie restriction with or without exercise on body composition and fat distribution. J Clin Endocrinol Metab 2007;92:865–872
12.Adams F. The Genuine Works of Hippocrates. Vol. 2 London: The Sydenham Society; 1849
13.Flatt J. Misconceptions in body weight regulation: Implications for the obesity pandemic. Crit Rev Clin Lab Sci 2012;49:150–165
14.Atwater WO. Food and Diet. Yearbook of Department of Agriculture 1894. Washington DC: Government Printing Office; 1895:357–388
15.Lighton JR. Measuring Metabolic Rates: A Manual for Scientists. New York: Oxford University Press; 2008
16.Webb P., Human Calorimeters. Endocrinology and Metabolism Series. Vol. 7 New York: Praeger; 1985
17.Atwater WO, Woods CD, Bryant AP. The Chemical Composition of American Food Materials. Washington, DC: Government Printing Office; 1906
18.Buchholz AC, Schoeller DA. Is a calorie a calorie? Am J Clin Nutr 2004;79:899S–906S
19.Moe PW. Future directions for energy requirements and food energy values. J Nutr 1994;124:1738S–1742S
20.Richardson HB. The respiratory quotient. Physiol Rev 1929;9:61–125
21.Hellerstein MK. No common energy currency: De novo lipogenesis as the road less traveled. Am J Clin Nutr 2001;74:707–708
22.Hellerstein MK. De novo lipogenesis in humans: Metabolic and regulatory aspects. Eur J Clin Nutr 1999;53:S53–S65
23.Hellerstein MK, Christiansen M, Kaempfer S, et al. Measurement of de novo hepatic lipogenesis in humans using stable isotopes. J Clin Invest 1991;87:1841–1852
24.Acheson KJ, Schutz Y, Bessard T, et al. Carbohydrate metabolism and de novo lipogenesis in human obesity. Am J Clin Nutr 1987;45:78–85
25.Diaz EO, Prentice AM, Goldberg GR, et al. Metabolic response to experimental overfeeding in lean and overweight healthy volunteers. Am J Clin Nutr 1992;56:641–655
26.Hill JO, Prentice AM. Sugar and body weight regulation. Am J Clin Nutr 1995;62:264S–273S
27.McDevitt RM, Bott SJ, Harding M, et al. De novo lipogenesis during controlled overfeeding with sucrose or glucose in lean and obese women. Am J Clin Nutr 2001;74:737–746
28.Harris JA, Benedict FG. A biometric study of human basal metabolism. Proc Natl Acad Sci USA 1918;4:370–373
29.Bouhlel E., Salhi Z., Bouhlel H., et al. Effect of Ramadan fasting on fuel oxidation during exercise in trained male rugby players. Diabetes Metab 2006;32:617–624
30.Bryson J, King S, Burns C, et al. Changes in glucose and lipid metabolism following weight loss produced by a very low calorie diet in obese subjects. Int J Obes Relat Metab Disord 1996;20:338–345
31.Valtueña S, Solà R, Salas-Salvadó J. A study of the prognostic respiratory markers of sustained weight loss in obese subjects after 28 days on VLCD. Int J Obes 1997;21:267–273
32.Finlayson G, Caudwell P, Gibbons C, et al. Low fat loss response after medium-term supervised exercise in obese is associated with exercise-induced increase in food reward. J Obes 2011;2011:pii.:615624.
33.Gropper S, Smith J. Advanced Nutrition and Human Metabolism. Belmont, CA: Wadsworth Cengage Learning; 2012
34.Cahill GF., Jr. Fuel metabolism in starvation. Annu Rev Nutr 2006;26:1–22
35.Felig P, Owen OE, Wahren J, et al. Amino acid metabolism during prolonged starvation. J Clin Invest 1969;48:584–594
36.McCue M. Specific dynamic action: A century of investigation. Comp Biochem Physiol A Mol Integr Physiol 2006;144:381–394
37.Westerterp KR. Diet induced thermogenesis. Nutr Metab 2004;1:5.
38.Jebb SA, Prentice AM, Goldberg GR, et al. Changes in macronutrient balance during over-and underfeeding assessed by 12-d continuous whole-body calorimetry. Am J Clin Nutr 1996;64:259–266
39.Prentice AM. Alcohol and obesity. Int J Obes 1995;19:S44–S50
40.Prentice AM. Are all calories equal? In: Cottrell R, ed. Weight Control. London: Chapman & Hall; 1995:8–33
41.Stubbs RJ, Harbron CG, Murgatroyd PR, et al. Covert manipulation of dietary fat and energy density: Effect on substrate flux and food intake in men eating ad libitum. Am J Clin Nutr 1995;62:316–329
42.Shetty PS, Prentice AM, Goldberg GR, et al. Alterations in fuel selection and voluntary food intake in response to isoenergetic manipulation of glycogen stores in humans. Am J Clin Nutr 1994;60:534–543
43.Prentice AM. Macronutrients as sources of food energy. Public Health Nutr 2005;8:932–939
44.McDevitt RM, Poppitt SD, Murgatroyd PR, et al. Macronutrient disposal during controlled overfeeding with glucose, fructose, sucrose, or fat in lean and obese women. Am J Clin Nutr 2000;72:369–377
45.Shelmet JJ, Reichard GA, Skutches CL, et al. Ethanol causes acute inhibition of carbohydrate, fat, and protein oxidation and insulin resistance. J Clin Invest 1988;81:1137–1145
46.Jessen C. Temperature regulation in humans and other mammals. Berlin Heidelberg: Springer-Verlag; 2012
47.Chen KY, Brychta RJ, Linderman JD, et al. Brown fat activation mediates cold-induced thermogenesis in adult humans in response to a mild decrease in ambient temperature. J Clin Endocrinol Metab 2013;98:E1218–E1223
48.Lee P, Smith S, Linderman J, et al. Temperature-acclimated brown adipose tissue modulates insulin sensitivity in humans. Diabetes 2014;63:3686–3698
49.Moellering DR, Smith DL., Jr.Ambient temperature and obesity. Curr Obes Rep 2012;1:26–34
50.Wijers SL, Schrauwen P, Saris WH, et al. Human skeletal muscle mitochondrial uncoupling is associated with cold induced adaptive thermogenesis. PLoS One 2008;3:e1777.
51.Chondronikola M, Volpi E, Børsheim E, et al. Brown adipose tissue activation is linked to distinct systemic effects on lipid metabolism in humans. Cell Metab 2016;23:1200–1206
52.Choi CS, Befroy DE, Codella R, et al. Paradoxical effects of increased expression of PGC-1α on muscle mitochondrial function and insulin-stimulated muscle glucose metabolism. Proc Natl Acad Sci USA 2008;105:19926–19931
53.Boström P, Wu J, Jedrychowski MP, et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012;481:463–468
54.Harms M, Seale P. Brown and beige fat: Development, function and therapeutic potential. Nat Med 2013;19:1252–1263
55.Tan DX, Manchester LC, Fuentes‐Broto L, et al. Significance and application of melatonin in the regulation of brown adipose tissue metabolism: Relation to human obesity. Obes Rev 2011;12:167–188
56.Cypess AM, Lehman S, Williams G, et al. Identification and importance of brown adipose tissue in adult humans. N Engl J Med 2009;360:1509–1517
57.van Marken Lichtenbelt WD, Vanhommerig JW, Smulders N., et al. Cold-activated brown adipose tissue in healthy men. N Engl J Med 2009;360:1500–1508
58.Handschin C, Spiegelman BM. Peroxisome proliferator-activated receptor γ coactivator 1 coactivators, energy homeostasis, and metabolism. Endocr Rev 2006;27:728–735
59.Valdés S, Maldonado‐Araque C, García‐Torres F, et al. Ambient temperature and prevalence of obesity in the Spanish population: The Di@bet.es study. Obesity 2014;22:2328–2332
60.Yang HK, Han K, Cho J-H, et al. Ambient temperature and prevalence of obesity: A nationwide population-based study in Korea. PLoS One 2015;10:e0141724.
61.Barber TM, Begbie H, Levy J. The incretin pathway as a new therapeutic target for obesity. Maturitas 2010;67:197–202
62.Kim W, Egan JM. The role of incretins in glucose homeostasis and diabetes treatment. Pharmacol Rev 2008;60:470–512
63.World Disasters Report 2011. Focus on Hunger and Malnutrition. In: Knight L. (ed). Geneva: International Federation of Red Cross and Red Crescent Societies; 2011: 251
64.Brandhorst S, Choi IY, Wei M, et al. , A periodic diet that mimics fasting promotes multi-system regeneration, enhanced cognitive performance, and healthspan. Cell Metab 2015;22:86–99
65.Schultz MB, Sinclair DA. When stem cells grow old: Phenotypes and mechanisms of stem cell aging. Development 2016;143:3–14
66.Guarente L. Calorie restriction and sirtuins revisited. Genes Dev 2013;27:2072–2085
67.Guarente L. The resurgence of NAD+. Science 2016;352:1396–1397
68.Pijl H. Longevity. The allostatic load of dietary restriction. Physiol Behav 2012;106:51–57
69.Muoio DM, Neufer PD. Lipid-induced mitochondrial stress and insulin action in muscle. Cell Metab 2012;15:595–605
70.Trepanowski JF, Canale RE, Marshall KE, et al. Impact of caloric and dietary restriction regimens on markers of health and longevity in humans and animals: A summary of available findings. Nutr J 2011;10:107.
71.Hall KD, Chen KY, Guo J, et al. , Energy expenditure and body composition changes after an isocaloric ketogenic diet in overweight and obese men. Am J Clin Nutr 2016;104:324–333
72.Weiss R, Bremer AA, Lustig RH. What is metabolic syndrome, and why are children getting it? Ann N Y Acad Sci 2013;1281:123–140
73.Bremer AA, Mietus-Snyder M, Lustig R.H. Toward a unifying hypothesis of metabolic syndrome. Pediatrics 2012;129:557–570
74.Prentice AM, Jebb SA. Obesity in Britain: Gluttony or sloth? BMJ 1995;311:437–439
75.Katz D. Unfattening our children: forks over feet. Int J Obes 2011;35:33–37
76.Weiss EP, Albert SG, Reeds DN, et al. Effects of matched weight loss from calorie restriction, exercise, or both on cardiovascular disease risk factors: A randomized intervention trial. Am J Clin Nutr 2016;104:576–586
77.Pontzer H, Raichlen DA, Wood BM, et al. Hunter-gatherer energetics and human obesity. PLoS One 2012;7:e40503.
78.Lefevre M, Redman LM, Heilbronn LK, et al. Caloric restriction alone and with exercise improves CVD risk in healthy non-obese individuals. Atherosclerosis 2009;203:206–213
79.Larson-Meyer DE, Heilbronn LK, Redman LM, et al. Effect of calorie restriction with or without exercise on insulin sensitivity, β-cell function, fat cell size, and ectopic lipid in overweight subjects. Diabetes Care 2006;29:1337–1344
80.Poirier P, Giles TD, Bray GA, et al. Obesity and cardiovascular disease pathophysiology, evaluation, and effect of weight loss. Arterioscler Thromb Vasc Biol 2006;26:968–976
81.Bastien M, Poirier P, Lemieux I, et al. Overview of epidemiology and contribution of obesity to cardiovascular disease. Prog Cardiovasc Dis 2014;56:369–381
82.Galassi A, Reynolds K, He J. Metabolic syndrome and risk of cardiovascular disease: A meta-analysis. Am J Med 2006;119:812–819
83.Bouhlel E, Shephard RJ. Optimizing Physical Performance During Fasting and Dietary Restriction: Implications for Athletes and Sports Medicine. Boca Raton, FL: CRC Press; 2015
84.Goldhamer AC, Klaper M, Foorohar A, et al. Water-only fasting and an exclusively plant foods diet in the management of stage IIIa, low-grade follicular lymphoma. BMJ Case Rep 2015:pii:
85.Safdie FM, Dorff T, Quinn D, et al. Fasting and cancer treatment in humans: A case series report. Aging (Albany NY) 2009;1:988–1007
86.Goldhamer AC, Lisle DJ, Sultana P, et al. , Medically supervised water-only fasting in the treatment of borderline hypertension. J Altern Complement Med 2002;8:643–650
87.Goldhamer AC. Initial cost of care results in medically supervised water-only fasting for treating high blood pressure and diabetes. J Altern Complement Med 2002;8:696–697
88.Stewart W, Fleming LW. Features of a successful therapeutic fast of 382 days’ duration. Postgrad Med J 1973;49:203–209
89.Eckel-Mahan KL, Patel VR, de Mateo S, et al. Reprogramming of the circadian clock by nutritional challenge. Cell 2013;155:1464–1478
90.Sahar S, Sassone-Corsi P. Regulation of metabolism: The circadian clock dictates the time. Trends Endocrinol Metab 2012;23:1–8
91.Dibner C, Schibler U, Albrecht U. The mammalian circadian timing system: Organization and coordination of central and peripheral clocks. Annu Rev Physiol 2010;72:517–549
92.Figueroa A, Hanzu F, Gomis R. Nutrition and the clock gene. Acta Endo 2015;11:489–491
93.Kantermann T, Juda M, Merrow M, et al. The human circadian clock’s seasonal adjustment is disrupted by daylight saving time. Curr Biol 2007;17:1996–2000
94.Merrow M, Spoelstra K, Roenneberg T. The circadian cycle: Daily rhythms from behaviour to genes. EMBO Rep 2005;6:930–935
95.Roenneberg T, Wirz-Justice A, Merrow M. Life between clocks: Daily temporal patterns of human chronotypes. J Biol Rhythms 2003;18:80–90
96.Prentice AM. Starvation in humans: Evolutionary background and contemporary implications. Mech Ageing Dev 2005;126:976–981
97.Ramis MR, Esteban S, Miralles A, et al. Caloric restriction, resveratrol and melatonin: Role of SIRT1 and implications for aging and related-diseases. Mech Ageing Dev 2015;146:28–41
98.Longo VD, Antebi A, Bartke A, et al. Interventions to slow aging in humans: Are we ready? Aging Cell 2015;14:497–510
99.Longo VD, Panda S. Fasting, circadian rhythms, and time-restricted feeding in healthy lifespan. Cell Metab 2016;23:1048–1059
100.Mercken EM, Carboneau BA, Krzysik-Walker SM, et al. Of mice and men: The benefits of caloric restriction, exercise, and mimetics. Ageing Res Rev 2012;11:390–398
101.Hubbard BP, Sinclair DA. Small molecule SIRT1 activators for the treatment of aging and age-related diseases. Trends Pharmacol Sci 2014;35:146–154
102.Lee BC, Kaya A, Gladyshev VN. Methionine restriction and life‐span control. Ann NY Acad Sci 2016;1363:116–124
103.McCarty MF, Barroso-Aranda J, Contreras F. The low-methionine content of vegan diets may make methionine restriction feasible as a life extension strategy. Med Hypotheses 2009;72:125–128
104.McIsaac RS, Lewis KN, Gibney PA, et al. From yeast to human: Exploring the comparative biology of methionine restriction in extending eukaryotic life span. Ann N Y Acad Sci 2016;1363:155–170

„Wolność nie jest więc przeciwieństwem determinizmu. Wolność to zdolność jednostki do uświadomienia sobie, że to ona jest zdeterminowana, do zatrzymania się między bodźcem a reakcją, a tym samym do opowiedzenia się, choćby w niewielkim stopniu, za jedną konkretną reakcją spośród kilku możliwych” – Rollo May

Najnowsze komentarze